I am writing the full report in another blog: Beginning of China ....
國際著名學術期刊《自然》最新發表一篇研究論文指出,包括日語、韓語、通古斯語、蒙古語、突厥語在內的阿爾泰語系(泛歐亞語系),可能起源於約9000年前的中國,並在農業的發展下傳播開來。因為古 DNA 高通量測序技術的出現和發展,古人類的研究取得了一系列突破性進展。研究揭示,該語系使用者是新石器時代生活在中國西遼河流域、種植小米的農民。隨著這些農民後來遷移到東北亞廣大地區,他們的語言向北向西流傳到西伯利亞和中亞草原,向東被帶到朝鮮半島和日本列島。現代漢語有著另一個源頭,漢藏語系大約與阿爾泰語系出現在同一時期,也與小米農耕相關,但地點在黃河流域。
Dec 15, 2022
古北歐亞人 Ancient North Eurasians (Ghost population)
在2012年的一個研究, 發現歐洲人和美洲土著的一個古基因交流(Ancient Genetic Admixture), 比歐洲人和東亞/東南亞人的更接近,這和“亞洲人到了美洲成為美洲土著”的說法衝突。 在2013年之後, 對一個發現於西伯利亞Balkai地區的2.4萬年前的小男孩遺骨的研究, 這個迷惑得到了解答, 這個小男孩被稱為MA-1(Mal'ta boy, 下面左上圖)。今天的歐洲人, 尤其是美洲土著的基因都和他有關聯 (比較下面左下圖基因的位置, 和右下圖MA-1的世界成分可知)。MA-1所屬的人群被稱為ANE(Ancient North Eurasians 古北亞歐人), 是典型的“Ghost 幽靈”人群,該人群不再以未混合的形式存在,他們幾乎與亞洲人和歐洲人的祖先, 同時在舊石器時期分離(下面右上圖)。這個ANE遺傳成分,主要向東傳播到美洲原住民,但也向西傳播到歐洲人, 以在西伯利亞發現的史前個體為代表。
大概在末次冰盛期(26.5 to 19~20 ka), 北歐大部、北美北部以及北亞的一部分均被大陸冰蓋覆蓋, ANE生活在亞歐大草原的廣袤範圍,以狩獵猛獁象、長毛犀牛和野牛為主, 橫跨大約7000公里,從黑海一直到西伯利亞。大約2萬年前,一部分ANE和東亞人混合,然後在大約1.5萬年前去了美洲, 美洲的土著的基因, 大約具有 14% 至 38% ANE成分(下面右下圖紅色)。(下面左上圖數字) 1, Oberkassel; 2, Hohle Fels; 3, Dolni Vestonice; 4, Kostenki-14. A Palaeolithic site with an individual belonging to mtDNA haplogroup B is represented by the square: 5, Tianyuan Cave. Notable Palaeolithic sites with Venus figurines are marked by brown circles: 6, Laussel; 7, Lespugue; 8, Grimaldi; 9, Willendorf; 10, Gargarino. Other notable Palaeolithic sites are shown by grey circles: 11, Sungir; 12, Yana RHS. (左下圖顯示) MA-1 is intermediate between modern western Eurasians and native Americans, but distant from east Asians.


在2013年的發表, 來自西伯利亞的舊石器時代晚期的基因組,證明了古北歐亞人(ANE)的存在,而且證明了他們攜帶男性單倍群 R*、歐洲單倍群 R1a 和 R1b 的祖先進化枝(Clade)。還發現了線粒體單倍群 U,這是中石器時代歐洲無處不在的母系譜系,在當代歐洲人中也很常見。從常染色體證據, 表明 MA-1 是現代西歐亞人的基礎,在遺傳上與現代美洲原住民密切相關,與東亞人沒有密切的親緣關係。對另一個可追溯到大約 17 ka 的西伯利亞中南部的 Afontova Gora-2 進行測序,揭示了與 MA-1 相似的常染色體遺傳特徵,表明該地區在整個末次盛冰期一直被人類占領。然而,早期的古北歐人也攜帶了東亞人的血統,比如田園洞人。 在西伯利亞東北Yana河 2019 年發現的 32.6 ka 樣本, 大約 22% 的貢獻來自早期東亞人。ANE在人類走出非洲演化樹, 看下面左圖。
隨著末次冰期的冰川消退,在全新世早期, 部分ANE人走入西歐亞部分地區, 也許是依靠獵殺猛獁象, 來獲取食物、衣服、建築材料以及象牙工具和武器的機會減少了。大約5000年前,在東歐大草原(Pontic-Caspian Steppe), 攜帶高ANE成分的東部狩獵採集者 (EHG) 和高加索狩獵採集者 (CHG)混合,形成了顏那亞(Yamnaya)文化, 約有 50% 的ANE成分。據信,他們的眼睛大部分是棕色的,淺色到中等膚色,棕色頭髮,有一些變化。 在2015 年的一項研究發現,顏那亞人在所有測試的古代種群中,對身高的計算遺傳選擇是最高的。之後從大約4800年前開始,他們大舉西遷,這就是歐洲人的第三個部分基因稱為草原基因 "Steppe DNA"(看下面右方右圖綠色), 可以看到北歐國家具有比較高ANE成分, 他們可能講的是一種非常古老的原始印歐語。


古希臘人相信有一群金髮碧眼的巨人,他們幸福地活到一千年,居住在遙遠北方的土地上,那裡日夜照耀。 他們是超級北方人,據信他們的家園位於巴爾幹半島、英國或烏拉爾以外的地方。遺傳學家David Reich認為,金色頭髮的 KITLG 基因是具有古北歐血統人群從歐亞草原大量攜帶進入歐洲的。已知最早的具有該等位基因的是來自 Afontova Gora 3 的西伯利亞中南部女性個體,其歷史可以追溯到1800年前。同樣地, 有2020年報告說該人群是使歐洲頭髮顏色變淺的原因(看下圖)。研究表明,現代歐洲人膚色較淺的兩個基因起源於大約 22000 至 28000 年前的中東和高加索地區,並在 8500 年前出現在安納托利亞,他們與新石器時代發展以及農業在歐洲的傳播有關。歐洲的另一波淺膚色人群與顏那亞文化和印歐人遷移有關,這些遷徙帶有古北歐亞血統。

A famous example of Tarim mummies with blonde hair is Princess of Xiaohe (塔里木盆地考古 Tarim Basin Archaeology) and was studied in below.

《自然》(Nature)2021-10-27 發表了塔里木盆地發現的青銅時代干屍屬於古北歐亞人(Ancient North Eurasian), 看下面的報告。
青銅時代新疆乾屍小河公主來自哪?中外團隊基因測序有新發現
澎湃新聞首席記者 賀梨萍 2021-10-27 23:00 來源:澎湃新聞
新疆塔里木的干屍“小河公主”流傳甚廣,小河墓地的發掘也曾被評為2004年度“中國十大考古新發現”之一。然而“小河公主”到底是哪里人卻一直是考古謎團。伊弟利斯·阿不都熱蘇勒和李文瑛在一篇文章中曾如此描述:從外觀看,小河墓地是一座橢圓形的沙山,高出地表7米餘,長74、寬35米。沙山表面密密叢叢矗立胡楊木柱百餘根,遠遠望去,猶如沙海中的一區叢林。
伊弟利斯·阿不都熱蘇勒為原新疆維吾爾自治區文物考古研究所所長、研究員,李文瑛當時為新疆維吾爾自治區文物考古研究所副所長、研究員,現任所長。他們所描述的小河墓地最早發現於上世紀三十年代,2000年後被再度探尋,最終引起國內外學術界的廣泛關注。然後,包括在塔里木盆地在內的新疆發現的這些史前乾屍,留給外界的謎團頗多。尤其是他們酷似白人的外表特徵、氈制和編織的羊毛服裝,以及可觀察到的農業畜牧經濟,包括牛、羊、小麥、大麥、小米,甚至是開菲爾(Kefir)奶酪,諸多的“西方”元素讓科學家們長期以來對其神秘起源感到困惑。
北京時間10月27日23時,頂級學術期刊《自然》(Nature)在線發表了由中德美韓四國團隊組成的合作團隊完成的研究,題為《青銅時代塔里木盆地干屍的基因組起源》(The genomic origins of the Bronze Age Tarim Basin mummies)。研究的通訊作者之一為吉林大學生命科學學院教授崔銀秋,李文瑛也是此項研究的作者。研究團隊分析了18具乾屍的基因組DNA:13具來自新疆南部塔里木盆地小河文化,可追溯至青銅時代中期(大約公元前2100-前1700年),5具來自新疆北部準噶爾盆地,可追溯至青銅時代早期(公元前3000-前2800年),這是首次對新疆地區史前人口進行基因組規模研究。研究團隊在論文中也提到,這些個體代表了該地區迄今為止挖掘出的最早的人類遺骸。
全基因組分析顯示,塔里木盆地發現的這些青銅時代乾屍屬於一支遺傳上獨立的本地人群。他們並不是外來者,而是曾經廣泛分佈的更新世群體的直系後代,而這些群體在上個冰河世紀末期基本消失了。這一群體被稱為古代北歐亞混血人(ANE),在當今人口的基因組中只存活了一小部分。這些發現否定了此前的種種假設,即這些乾屍來自從現今西伯利亞、阿富汗北部或中亞山區遷徙而來的人群後代。 《自然》還同期發表了由納扎爾巴耶夫大學人類學和社會學系的Paula N. Doumani Dupuy撰寫的新聞與觀點文章中,Dupuy進一步探討了本文的關鍵發現,及其對亞洲內陸史前史的意義。她認為,研究團隊“回答了小河文化的遺傳起源。現在要靠學者們努力合作,進一步解釋界定了亞洲內陸青銅時代的動態及多樣化的文化交流模式”。
東西文化交匯、亞歐重要樞紐
新疆作為絲綢之路的一部分,地處東西方文化的交匯處,長期以來一直是亞歐各國人民、文化、農業、語言交流的重要樞紐。東西走向的天山山脈橫貫新疆的東西,分隔出了地理上的北疆和南疆。北疆有準噶爾盆地,南疆則包括塔里木盆地。研究團隊在論文中寫道,北疆的準噶爾盆地的中央是古爾班通古特沙漠(Gurbantünggüt),它的周圍是大片草原,傳統上居住著游牧民族。南疆的塔里木盆地是一個乾燥的內海,現在形成了塔克拉瑪干沙漠。雖然大部分地區不適宜居住,但塔里木盆地也有小綠洲和河流,這些是由周圍高山融化的冰川和積雪形成的。
值得注意的是,相比古埃及精心製作的木乃伊,新疆的干屍均是自然形成。一位考古領域的專家對澎湃新聞(www.thepaper.cn)記者表示,“新疆的干屍很多,各個時期的都有,主要是氣候乾燥造成的。” 論文提到,在塔里木盆地各處發現的干屍中,最早的是古墓溝墓地(公元前2135-前1939年)、小河墓地(公元前1884-前1736年)和北方墓地(公元前1785-前1664年)墓葬底層發現的干屍。這些和相關的青銅器時代遺址基於其共同的物質文化被歸類到小河考古範圍內。
小河墓地俯瞰圖; 小河墓地代表性的帶槳船棺,棺上蓋著牛皮; 小河墓地M11號墓葬中一個自然保存的女性乾屍:



從考古的角度看,小河乾屍中“西方元素”的起源,一直是全球學術界的焦點,學者們也提出了多種假設。論文中羅列了包括顏那亞人/阿凡納謝沃人(Yamnaya/Afanasievo)草原假說、綠洲假說和亞洲內陸山脈走廊(IAMC)島嶼生物地理學假說。
第一種假說認為,阿爾泰-薩揚山脈中與阿凡納謝沃人相關的青銅時代早期種群通過準噶爾盆地傳播到塔里木盆地,隨後在公元前2000年左右建立了農業畜牧群落。第二種假說認為,塔里木盆地最初由從阿富汗、土庫曼斯坦和烏茲別克斯坦的沙漠綠洲農民(BMAC)遷移而來,對這一假設的支持很大程度上是基於這兩個地區的農業和灌溉系統的相似性,反映了對沙漠環境的適應,以及這兩個地區儀式上使用麻黃的證據。IAMC島嶼生物地理學假說則假設了小河最初人口來自中亞山區。
然而,作者們在論文中指出,最近的考古基因組研究表明,西伯利亞南部的青銅時代的阿凡納謝沃人和IAMC/BMAC群體具有可區分的遺傳特徵,這些特徵也不同於亞洲內陸的農業畜牧前狩獵-採集群體。因此,他們認為,對新疆青銅時代人口的考古基因組研究,為重建準噶爾盆地和塔里木盆地的人口歷史和青銅時代小河區域的起源提供了有力的途徑。
基因上是獨立的,但文化上是世界性的
為了更好地理解塔里木盆地干屍的起源和早期人口,研究團隊作者分析了在新疆發現的18具乾屍的DNA:13具來自新疆南部塔里木盆地小河文化,可追溯至青銅時代中期(公元前大約2100-1700年),5具來自新疆北部準噶爾盆地,可追溯至青銅時代早期(公元前3000-2800年)。他們還檢測了7具塔里木盆地干屍牙齒上礦化斑塊中的蛋白質,以重建他們的飲食。
新疆青铜器时代考古遗址概况:
 令人驚訝的是,研究團隊發現他們的分析結果並不符合上述三種假說。塔里木盆地的干屍根本不是外來者,而是曾經廣泛分佈的更新世群體的直系後代,這些群體在上個冰河世紀末期基本消失了。這一群體被稱為古北歐亞人(ANE),在當今人口的基因組中只存活了一小部分,西伯利亞和美洲的土著人口擁有已知的最高比例,約為40%。研究認為,與今天的人群相比,塔里木盆地的干屍沒有顯示出與任何其他全新世群體混合的證據,而是形成了以前未知的基因隔離,在定居塔里木盆地之前可能經歷了一個極端和長期的基因瓶頸。
令人驚訝的是,研究團隊發現他們的分析結果並不符合上述三種假說。塔里木盆地的干屍根本不是外來者,而是曾經廣泛分佈的更新世群體的直系後代,這些群體在上個冰河世紀末期基本消失了。這一群體被稱為古北歐亞人(ANE),在當今人口的基因組中只存活了一小部分,西伯利亞和美洲的土著人口擁有已知的最高比例,約為40%。研究認為,與今天的人群相比,塔里木盆地的干屍沒有顯示出與任何其他全新世群體混合的證據,而是形成了以前未知的基因隔離,在定居塔里木盆地之前可能經歷了一個極端和長期的基因瓶頸。
“為了更好地了解歐亞大陸內部的遺傳歷史,考古學家長期以來一直在尋找全新世ANE族,我們在最意想不到的地方發現了一個,”研究通訊作者之一、首爾大學生命科學學院教授Choongwon Jeong表示。值得關注的是,與塔里木盆地不同的是,相鄰的準噶爾盆地最早的居民不僅是當地居民的後代,也是西方草原牧民的後代,即阿凡納謝沃人(Afanasievo),這是一個與早期青銅時代的顏那亞人(Yamanya)有著強烈遺傳聯繫的游牧群體。研究團隊認為,這些發現也增加了我們對顏那亞人祖先向東傳播的理解,以及當他們第一次遇到亞洲內部的人口時發生基因混合的場景。
研究同時強調,雖然塔里木盆地的群體在基因上是隔離獨立的,但他們在文化上並非如此。對他們牙石的蛋白質組學分析證實,牛、綿羊和山羊的牧場已經被當時的人口所實踐,他們也很清楚周圍不同的文化、烹飪方法和技術。“儘管在基因上是孤立的,但塔里木盆地的青銅時代的人們在文化上是非常國際化的——他們以西亞的小麥和奶製品、東亞的小米和中亞的麻黃等藥用植物為食。”該研究的通訊作者之一、哈佛大學人類學教授、MPI-EVA研究小組負責人Christina Warinner博士說。
論文中提到,綜合考慮這些發現可以推測,小河文化當時的創始人口非常好地意識到了塔里木盆地之外的不同的技術和文化,他們發展了獨特的文化。“重建塔里木盆地干屍的起源對我們對該地區的理解產生了變革性的影響,我們將繼續研究其他時代的古代人類基因組,以更深入地了解歐亞大草原上的人類遷移歷史。”該研究另一通訊作者、吉林大學生命科學學院教授崔銀秋補充道。
Dupuy在新聞與觀點文章的最後也寫道,南疆和北疆這兩個截然不同的故事表明,即使是對古代DNA最全面的分析,也無法完全揭示有助於考古的文化實踐和選擇。她認為這項最新的研究對塔里木盆地獨特而國際化的小河文化以及準噶爾盆地人口的描述,為先前的觀點提供了一個例子,即文化思想和新技術在亞洲內部流動,並進行各種迭代。
吳偉榮2021秋
大概在末次冰盛期(26.5 to 19~20 ka), 北歐大部、北美北部以及北亞的一部分均被大陸冰蓋覆蓋, ANE生活在亞歐大草原的廣袤範圍,以狩獵猛獁象、長毛犀牛和野牛為主, 橫跨大約7000公里,從黑海一直到西伯利亞。大約2萬年前,一部分ANE和東亞人混合,然後在大約1.5萬年前去了美洲, 美洲的土著的基因, 大約具有 14% 至 38% ANE成分(下面右下圖紅色)。(下面左上圖數字) 1, Oberkassel; 2, Hohle Fels; 3, Dolni Vestonice; 4, Kostenki-14. A Palaeolithic site with an individual belonging to mtDNA haplogroup B is represented by the square: 5, Tianyuan Cave. Notable Palaeolithic sites with Venus figurines are marked by brown circles: 6, Laussel; 7, Lespugue; 8, Grimaldi; 9, Willendorf; 10, Gargarino. Other notable Palaeolithic sites are shown by grey circles: 11, Sungir; 12, Yana RHS. (左下圖顯示) MA-1 is intermediate between modern western Eurasians and native Americans, but distant from east Asians.
在2013年的發表, 來自西伯利亞的舊石器時代晚期的基因組,證明了古北歐亞人(ANE)的存在,而且證明了他們攜帶男性單倍群 R*、歐洲單倍群 R1a 和 R1b 的祖先進化枝(Clade)。還發現了線粒體單倍群 U,這是中石器時代歐洲無處不在的母系譜系,在當代歐洲人中也很常見。從常染色體證據, 表明 MA-1 是現代西歐亞人的基礎,在遺傳上與現代美洲原住民密切相關,與東亞人沒有密切的親緣關係。對另一個可追溯到大約 17 ka 的西伯利亞中南部的 Afontova Gora-2 進行測序,揭示了與 MA-1 相似的常染色體遺傳特徵,表明該地區在整個末次盛冰期一直被人類占領。然而,早期的古北歐人也攜帶了東亞人的血統,比如田園洞人。 在西伯利亞東北Yana河 2019 年發現的 32.6 ka 樣本, 大約 22% 的貢獻來自早期東亞人。ANE在人類走出非洲演化樹, 看下面左圖。
隨著末次冰期的冰川消退,在全新世早期, 部分ANE人走入西歐亞部分地區, 也許是依靠獵殺猛獁象, 來獲取食物、衣服、建築材料以及象牙工具和武器的機會減少了。大約5000年前,在東歐大草原(Pontic-Caspian Steppe), 攜帶高ANE成分的東部狩獵採集者 (EHG) 和高加索狩獵採集者 (CHG)混合,形成了顏那亞(Yamnaya)文化, 約有 50% 的ANE成分。據信,他們的眼睛大部分是棕色的,淺色到中等膚色,棕色頭髮,有一些變化。 在2015 年的一項研究發現,顏那亞人在所有測試的古代種群中,對身高的計算遺傳選擇是最高的。之後從大約4800年前開始,他們大舉西遷,這就是歐洲人的第三個部分基因稱為草原基因 "Steppe DNA"(看下面右方右圖綠色), 可以看到北歐國家具有比較高ANE成分, 他們可能講的是一種非常古老的原始印歐語。
古希臘人相信有一群金髮碧眼的巨人,他們幸福地活到一千年,居住在遙遠北方的土地上,那裡日夜照耀。 他們是超級北方人,據信他們的家園位於巴爾幹半島、英國或烏拉爾以外的地方。遺傳學家David Reich認為,金色頭髮的 KITLG 基因是具有古北歐血統人群從歐亞草原大量攜帶進入歐洲的。已知最早的具有該等位基因的是來自 Afontova Gora 3 的西伯利亞中南部女性個體,其歷史可以追溯到1800年前。同樣地, 有2020年報告說該人群是使歐洲頭髮顏色變淺的原因(看下圖)。研究表明,現代歐洲人膚色較淺的兩個基因起源於大約 22000 至 28000 年前的中東和高加索地區,並在 8500 年前出現在安納托利亞,他們與新石器時代發展以及農業在歐洲的傳播有關。歐洲的另一波淺膚色人群與顏那亞文化和印歐人遷移有關,這些遷徙帶有古北歐亞血統。
A famous example of Tarim mummies with blonde hair is Princess of Xiaohe (塔里木盆地考古 Tarim Basin Archaeology) and was studied in below.
《自然》(Nature)2021-10-27 發表了塔里木盆地發現的青銅時代干屍屬於古北歐亞人(Ancient North Eurasian), 看下面的報告。
青銅時代新疆乾屍小河公主來自哪?中外團隊基因測序有新發現
澎湃新聞首席記者 賀梨萍 2021-10-27 23:00 來源:澎湃新聞
新疆塔里木的干屍“小河公主”流傳甚廣,小河墓地的發掘也曾被評為2004年度“中國十大考古新發現”之一。然而“小河公主”到底是哪里人卻一直是考古謎團。伊弟利斯·阿不都熱蘇勒和李文瑛在一篇文章中曾如此描述:從外觀看,小河墓地是一座橢圓形的沙山,高出地表7米餘,長74、寬35米。沙山表面密密叢叢矗立胡楊木柱百餘根,遠遠望去,猶如沙海中的一區叢林。
伊弟利斯·阿不都熱蘇勒為原新疆維吾爾自治區文物考古研究所所長、研究員,李文瑛當時為新疆維吾爾自治區文物考古研究所副所長、研究員,現任所長。他們所描述的小河墓地最早發現於上世紀三十年代,2000年後被再度探尋,最終引起國內外學術界的廣泛關注。然後,包括在塔里木盆地在內的新疆發現的這些史前乾屍,留給外界的謎團頗多。尤其是他們酷似白人的外表特徵、氈制和編織的羊毛服裝,以及可觀察到的農業畜牧經濟,包括牛、羊、小麥、大麥、小米,甚至是開菲爾(Kefir)奶酪,諸多的“西方”元素讓科學家們長期以來對其神秘起源感到困惑。
北京時間10月27日23時,頂級學術期刊《自然》(Nature)在線發表了由中德美韓四國團隊組成的合作團隊完成的研究,題為《青銅時代塔里木盆地干屍的基因組起源》(The genomic origins of the Bronze Age Tarim Basin mummies)。研究的通訊作者之一為吉林大學生命科學學院教授崔銀秋,李文瑛也是此項研究的作者。研究團隊分析了18具乾屍的基因組DNA:13具來自新疆南部塔里木盆地小河文化,可追溯至青銅時代中期(大約公元前2100-前1700年),5具來自新疆北部準噶爾盆地,可追溯至青銅時代早期(公元前3000-前2800年),這是首次對新疆地區史前人口進行基因組規模研究。研究團隊在論文中也提到,這些個體代表了該地區迄今為止挖掘出的最早的人類遺骸。
全基因組分析顯示,塔里木盆地發現的這些青銅時代乾屍屬於一支遺傳上獨立的本地人群。他們並不是外來者,而是曾經廣泛分佈的更新世群體的直系後代,而這些群體在上個冰河世紀末期基本消失了。這一群體被稱為古代北歐亞
東西文化交匯、亞歐重要樞紐
新疆作為絲綢之路的一部分,地處東西方文化的交匯處,長期以來一直是亞歐各國人民、文化、農業、語言交流的重要樞紐。東西走向的天山山脈橫貫新疆的東西,分隔出了地理上的北疆和南疆。北疆有準噶爾盆地,南疆則包括塔里木盆地。研究團隊在論文中寫道,北疆的準噶爾盆地的中央是古爾班通古特沙漠(Gurbantünggüt),它的周圍是大片草原,傳統上居住著游牧民族。南疆的塔里木盆地是一個乾燥的內海,現在形成了塔克拉瑪干沙漠。雖然大部分地區不適宜居住,但塔里木盆地也有小綠洲和河流,這些是由周圍高山融化的冰川和積雪形成的。
值得注意的是,相比古埃及精心製作的木乃伊,新疆的干屍均是自然形成。一位考古領域的專家對澎湃新聞(www.thepaper.cn)記者表示,“新疆的干屍很多,各個時期的都有,主要是氣候乾燥造成的。” 論文提到,在塔里木盆地各處發現的干屍中,最早的是古墓溝墓地(公元前2135-前1939年)、小河墓地(公元前1884-前1736年)和北方墓地(公元前1785-前1664年)墓葬底層發現的干屍。這些和相關的青銅器時代遺址基於其共同的物質文化被歸類到小河考古範圍內。
小河墓地俯瞰圖; 小河墓地代表性的帶槳船棺,棺上蓋著牛皮; 小河墓地M11號墓葬中一個自然保存的女性乾屍:
從考古的角度看,小河乾屍中“西方元素”的起源,一直是全球學術界的焦點,學者們也提出了多種假設。論文中羅列了包括顏那亞人/阿凡納謝沃人(Yamnaya/Afanasievo)草原假說、綠洲假說和亞洲內陸山脈走廊(IAMC)島嶼生物地理學假說。
第一種假說認為,阿爾泰-薩揚山脈中與阿凡納謝沃人相關的青銅時代早期種群通過準噶爾盆地傳播到塔里木盆地,隨後在公元前2000年左右建立了農業畜牧群落。第二種假說認為,塔里木盆地最初由從阿富汗、土庫曼斯坦和烏茲別克斯坦的沙漠綠洲農民(BMAC)遷移而來,對這一假設的支持很大程度上是基於這兩個地區的農業和灌溉系統的相似性,反映了對沙漠環境的適應,以及這兩個地區儀式上使用麻黃的證據。IAMC島嶼生物地理學假說則假設了小河最初人口來自中亞山區。
然而,作者們在論文中指出,最近的考古基因組研究表明,西伯利亞南部的青銅時代的阿凡納謝沃人和IAMC/BMAC群體具有可區分的遺傳特徵,這些特徵也不同於亞洲內陸的農業畜牧前狩獵-採集群體。因此,他們認為,對新疆青銅時代人口的考古基因組研究,為重建準噶爾盆地和塔里木盆地的人口歷史和青銅時代小河區域的起源提供了有力的途徑。
基因上是獨立的,但文化上是世界性的
為了更好地理解塔里木盆地干屍的起源和早期人口,研究團隊作者分析了在新疆發現的18具乾屍的DNA:13具來自新疆南部塔里木盆地小河文化,可追溯至青銅時代中期(公元前大約2100-1700年),5具來自新疆北部準噶爾盆地,可追溯至青銅時代早期(公元前3000-2800年)。他們還檢測了7具塔里木盆地干屍牙齒上礦化斑塊中的蛋白質,以重建他們的飲食。
新疆青铜器时代考古遗址概况:
令人驚訝的是,研究團隊發現他們的分析結果並不符合上述三種假說。塔里木盆地的干屍根本不是外來者,而是曾經廣泛分佈的更新世群體的直系後代,這些群體在上個冰河世紀末期基本消失了。這一群體被稱為古北歐亞人(ANE),在當今人口的基因組中只存活了一小部分,西伯利亞和美洲的土著人口擁有已知的最高比例,約為40%。研究認為,與今天的人群相比,塔里木盆地的干屍沒有顯示出與任何其他全新世群體混合的證據,而是形成了以前未知的基因隔離,在定居塔里木盆地之前可能經歷了一個極端和長期的基因瓶頸。“為了更好地了解歐亞大陸內部的遺傳歷史,考古學家長期以來一直在尋找全新世ANE族,我們在最意想不到的地方發現了一個,”研究通訊作者之一、首爾大學生命科學學院教授Choongwon Jeong表示。值得關注的是,與塔里木盆地不同的是,相鄰的準噶爾盆地最早的居民不僅是當地居民的後代,也是西方草原牧民的後代,即阿凡納謝沃人(Afanasievo),這是一個與早期青銅時代的顏那亞人(Yamanya)有著強烈遺傳聯繫的游牧群體。研究團隊認為,這些發現也增加了我們對顏那亞人祖先向東傳播的理解,以及當他們第一次遇到亞洲內部的人口時發生基因混合的場景。
研究同時強調,雖然塔里木盆地的群體在基因上是隔離獨立的,但他們在文化上並非如此。對他們牙石的蛋白質組學分析證實,牛、綿羊和山羊的牧場已經被當時的人口所實踐,他們也很清楚周圍不同的文化、烹飪方法和技術。“儘管在基因上是孤立的,但塔里木盆地的青銅時代的人們在文化上是非常國際化的——他們以西亞的小麥和奶製品、東亞的小米和中亞的麻黃等藥用植物為食。”該研究的通訊作者之一、哈佛大學人類學教授、MPI-EVA研究小組負責人Christina Warinner博士說。
論文中提到,綜合考慮這些發現可以推測,小河文化當時的創始人口非常好地意識到了塔里木盆地之外的不同的技術和文化,他們發展了獨特的文化。“重建塔里木盆地干屍的起源對我們對該地區的理解產生了變革性的影響,我們將繼續研究其他時代的古代人類基因組,以更深入地了解歐亞大草原上的人類遷移歷史。”該研究另一通訊作者、吉林大學生命科學學院教授崔銀秋補充道。
Dupuy在新聞與觀點文章的最後也寫道,南疆和北疆這兩個截然不同的故事表明,即使是對古代DNA最全面的分析,也無法完全揭示有助於考古的文化實踐和選擇。她認為這項最新的研究對塔里木盆地獨特而國際化的小河文化以及準噶爾盆地人口的描述,為先前的觀點提供了一個例子,即文化思想和新技術在亞洲內部流動,並進行各種迭代。
吳偉榮2021秋
Dec 19, 2021
早期遷徙人類留在或離開非洲 Early Human Migrations stayed in or out of Africa
(2022.2.15) 更多的了解關於早期遷徙人類 More understanding on early human migrations.
請看下面在最後, 留在或離開非洲遷徙人類的現代人類後代圖片。
Please see below at the end, the pictures of modern descendants of early humans stayed in or out of Africa.
(Written 2019.6) Original Title: 早期人類沿海遷徙 Early Human Coastal Migrations:
在過去的幾年裡,基因檢測的進步改變了我們對早期歷史和文明的看法, 也改變了我們在旅行時了解當地人和文化的方式。使用單倍群不是政治和種族性的,因為它不是種族的標記,而是指出人類的起源。 現在英文維基百科,都有加這些資料。 In the past few years, the advance of genetic testing changes the way we look at early history and civilization. This is also change the ways we understand local people and culture when traveling. Using haplogroup is not political and racial related because it is not an identifier of ethnicity but indicates the origin of a population.
下面地圖顯示現代人的單倍群CF和DE,他們是M168的後代,部分大約7萬年前離開了非洲。
CF離開後再也沒有回來了, 然而DE分裂成D和E,E留在非洲成為Negroid大黑人(E1b1a),其中班圖人佔據了大部分撒哈拉以南地區,成為非洲最大的族群(76%)。 另一族群(E1b1b)被視為高加索種, 佔據了北非和非洲之角(17%)。Below map shows modern humans of haplogroups CF and DE, who were M168's offsprings, left Africa about 70,000 years ago. CF left and never returned. Then DE splitted into D and E. E stayed in Africa to become Negroid (E1b1a) in which Bantu occuppied most sub-Sahara and became the biggest group in Africa (76%). Another group (E1b1b), considered as Caucasiod, occuppied North and Horn of Africa (17%).
DE分裂後,據相信D先進入亞洲大陸,成為第一個現代人類定居東亞大陸者。他們成了羌族,白馬族和藏族等。 後來去了東南亞成為Negritos小黑人, 然後到了日本成為阿伊努人。 D-人群僅佔世界人口的2%。 請看下面現代人圖片, 顯示他們的多樣性。 After DE split, it is believed D went to mainland Asia first and became first human settlers in east Asia mainland.They became Qiang, Baima (means white horse) and Tibetans etc. After they went southeast Asia to become Negritos (means little blacks), and to Japan to become Ainu people. D-people only occupy 2% of world population. Please see below modern human pictures showing their diversity.
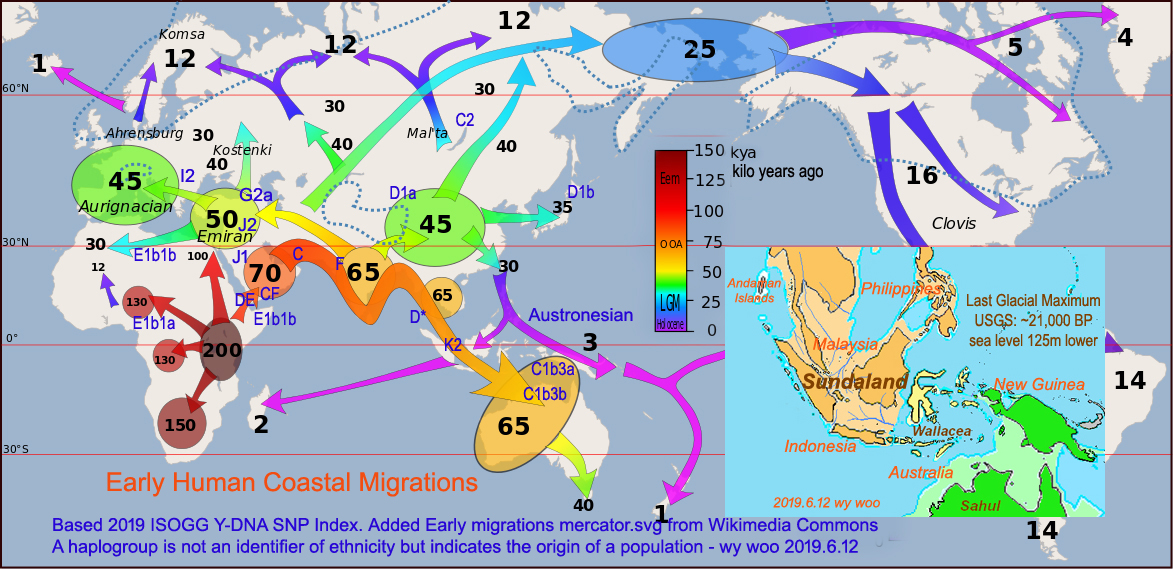
人類走出非洲(Out of Africa): CT-M168→DE,CF→C,F→G,HIJK→H,IJK→IJK→IJ,K→LT,K2→K2a(→NO),K2b(→MS,P→Q,R)
Note: 幾年前我在Previous Posts寫: "在7萬多年前人類走出非洲,其中D和E型的人群可能是在六、七萬年前紅海附近分離。攜帶E型的人回到非洲,成為非洲西部的Negroid大黑人;而攜帶D型的人輾轉向東遷徙,成為東南亞的Negrito小黑人"。 最近有人認為, 走出非洲不是七萬年前的M168的時候,DE和E並沒有走出, 而是更晚的6万年之內,C D F三支分別走出。
我們在摩洛哥旅行之後, 我發現E1b1b = Hamites(含族)不是黑人。
我們在巴爾幹半島和希臘旅行之後, 我發現希臘大陸人起源更接近北非E1b1b,而Create島人則來自西亞J2。
大約3600年前,猛烈火山爆發摧毀了希臘的Santorini島,留下了幾百米厚的火山灰。 Akrotiri鎮像Pompeii一樣被埋葬,為現代考古學家保留了許多精美的壁畫和藝術品。 請看史前人類和住宅視頻。
中國官方2018公佈"中華文明探源工程"成果, 距今約5,000年前,良著文化最早進入文明社會, 距今3,800年, 中華文明在自身發展過程中,廣泛吸收了外來文明的影響。源自西亞,中亞等地區的小麥栽培技術,黃牛和綿羊等家畜的飼養以及青銅冶煉技術逐步融入中華文明之中。
The new Y-DNA Haplogroup Trees 單倍群樹 (2022-2-14 Wikipedia) below show haplogroups A,B,E stayed in and haplogroups C,D,F out of Africa. Map notations new classification: D1a1a-M15, D1a1b-P99, D1a2a-M55, D1a2b-Y34637

上面地圖 Map Notations: (Written 2019.6)
C1b3a (formerly C2-M38): Indonesia, New Guinea, Melanesia, Micronesia, Polynesia
C1b3b (formerly C4-M347): Indigenous Australians
C2 (formerly C3-M217): Mongols, Kazakhs, Tungusic, Paleosiberians, Na-Dene
D* (M174): Negritos of Andaman Islands & Southesat Asia
D1a1 (formerly D1-M15): Qiang (羌族), Tibetan,
D1a2 (formerly D3-P99): Baima (白馬族), Pumi (普米族),
D1b (formerly D2-M55): Ainu, Yamato Japanese, Ryukyuan (Okinawa)
E1b1a (V38): Sub-Sahara, Negroid - Niger Congo (Bantu, Hutu, Tutsi, 76% of Africa)
E1b1b (M215): North Africa, Horn of Africa, Hamites (含族, Berber, Cushitic, Nilotic-Maasai, , 17% of Africa)
K2: Ancestor of most Asians, Europeans and Americans, 60% of world population
I2: Early Europeans (Balkans, Sardinia) before LGM (Last Glacial Maximum)
J1 (M267): Arabian Peninsula, Sudan, Ethiopia, West Asia, North Africa, Semites (閃族, Arabs & Jews etc.)
J2 (M172): Ancient Near East, Italy, Mediterranean littoral, Central Asia, South Asia
G2a (P15): Caucasia, Sardinia, Central & Southern Italy, Neolithic farmers & herders 6-9 kya. Ötzi the Iceman G2a2a2 (L91) 5 kya
Notable large expansions and migrations in early history:
Indo-European migrations 6 kya from Eurasian Pontic-Caspian steppe
Astronesin migrations 5 kya from Taiwan
Bantu expansions 4 kya from Western Africa
Arab conquests 7th century from Arabian Peninsula
Below pictures show the modern descendants of haplogroups A,B,C,D,E without F which constitutes most paternal lineages outside of Africa. Some pictures were taken by me and others were downloaded from internet. The picks were based upon the ethnic aspects of the humans. Fig (1): Modern descendants of Haplogroups C,D stayed out of Africa 單倍群C,D離開非洲的現代人類後代:

Fig (2): Modern descendants of Haplogroups A,B,E stayed in Africa 單倍群A,B,E留在非洲的現代人類後代:

吳偉榮2019夏 - revived 2022冬春
請看下面在最後, 留在或離開非洲遷徙人類的現代人類後代圖片。
Please see below at the end, the pictures of modern descendants of early humans stayed in or out of Africa.
(Written 2019.6) Original Title: 早期人類沿海遷徙 Early Human Coastal Migrations:
在過去的幾年裡,基因檢測的進步改變了我們對早期歷史和文明的看法, 也改變了我們在旅行時了解當地人和文化的方式。使用單倍群不是政治和種族性的,因為它不是種族的標記,而是指出人類的起源。 現在英文維基百科,都有加這些資料。 In the past few years, the advance of genetic testing changes the way we look at early history and civilization. This is also change the ways we understand local people and culture when traveling. Using haplogroup is not political and racial related because it is not an identifier of ethnicity but indicates the origin of a population.
下面地圖顯示現代人的單倍群CF和DE,他們是M168的後代,部分大約7萬年前離開了非洲。
CF離開後再也沒有回來了, 然而DE分裂成D和E,E留在非洲成為Negroid大黑人(E1b1a),其中班圖人佔據了大部分撒哈拉以南地區,成為非洲最大的族群(76%)。 另一族群(E1b1b)被視為高加索種, 佔據了北非和非洲之角(17%)。Below map shows modern humans of haplogroups CF and DE, who were M168's offsprings, left Africa about 70,000 years ago. CF left and never returned. Then DE splitted into D and E. E stayed in Africa to become Negroid (E1b1a) in which Bantu occuppied most sub-Sahara and became the biggest group in Africa (76%). Another group (E1b1b), considered as Caucasiod, occuppied North and Horn of Africa (17%).
DE分裂後,據相信D先進入亞洲大陸,成為第一個現代人類定居東亞大陸者。他們成了羌族,白馬族和藏族等。 後來去了東南亞成為Negritos小黑人, 然後到了日本成為阿伊努人。 D-人群僅佔世界人口的2%。 請看下面現代人圖片, 顯示他們的多樣性。 After DE split, it is believed D went to mainland Asia first and became first human settlers in east Asia mainland.They became Qiang, Baima (means white horse) and Tibetans etc. After they went southeast Asia to become Negritos (means little blacks), and to Japan to become Ainu people. D-people only occupy 2% of world population. Please see below modern human pictures showing their diversity.
人類走出非洲(Out of Africa): CT-M168→DE,CF→C,F→G,HIJK→H,IJK→IJK→IJ,K→LT,K2→K2a(→NO),K2b(→MS,P→Q,R)
Note: 幾年前我在Previous Posts寫: "在7萬多年前人類走出非洲,其中D和E型的人群可能是在六、七萬年前紅海附近分離。攜帶E型的人回到非洲,成為非洲西部的Negroid大黑人;而攜帶D型的人輾轉向東遷徙,成為東南亞的Negrito小黑人"。 最近有人認為, 走出非洲不是七萬年前的M168的時候,DE和E並沒有走出, 而是更晚的6万年之內,C D F三支分別走出。
我們在摩洛哥旅行之後, 我發現E1b1b = Hamites(含族)不是黑人。
我們在巴爾幹半島和希臘旅行之後, 我發現希臘大陸人起源更接近北非E1b1b,而Create島人則來自西亞J2。
大約3600年前,猛烈火山爆發摧毀了希臘的Santorini島,留下了幾百米厚的火山灰。 Akrotiri鎮像Pompeii一樣被埋葬,為現代考古學家保留了許多精美的壁畫和藝術品。 請看史前人類和住宅視頻。
中國官方2018公佈"中華文明探源工程"成果, 距今約5,000年前,良著文化最早進入文明社會, 距今3,800年, 中華文明在自身發展過程中,廣泛吸收了外來文明的影響。源自西亞,中亞等地區的小麥栽培技術,黃牛和綿羊等家畜的飼養以及青銅冶煉技術逐步融入中華文明之中。
The new Y-DNA Haplogroup Trees 單倍群樹 (2022-2-14 Wikipedia) below show haplogroups A,B,E stayed in and haplogroups C,D,F out of Africa. Map notations new classification: D1a1a-M15, D1a1b-P99, D1a2a-M55, D1a2b-Y34637
上面地圖 Map Notations: (Written 2019.6)
C1b3a (formerly C2-M38): Indonesia, New Guinea, Melanesia, Micronesia, Polynesia
C1b3b (formerly C4-M347): Indigenous Australians
C2 (formerly C3-M217): Mongols, Kazakhs, Tungusic, Paleosiberians, Na-Dene
D* (M174): Negritos of Andaman Islands & Southesat Asia
D1a1 (formerly D1-M15): Qiang (羌族), Tibetan,
D1a2 (formerly D3-P99): Baima (白馬族), Pumi (普米族),
D1b (formerly D2-M55): Ainu, Yamato Japanese, Ryukyuan (Okinawa)
E1b1a (V38): Sub-Sahara, Negroid - Niger Congo (Bantu, Hutu, Tutsi, 76% of Africa)
E1b1b (M215): North Africa, Horn of Africa, Hamites (含族, Berber, Cushitic, Nilotic-Maasai, , 17% of Africa)
K2: Ancestor of most Asians, Europeans and Americans, 60% of world population
I2: Early Europeans (Balkans, Sardinia) before LGM (Last Glacial Maximum)
J1 (M267): Arabian Peninsula, Sudan, Ethiopia, West Asia, North Africa, Semites (閃族, Arabs & Jews etc.)
J2 (M172): Ancient Near East, Italy, Mediterranean littoral, Central Asia, South Asia
G2a (P15): Caucasia, Sardinia, Central & Southern Italy, Neolithic farmers & herders 6-9 kya. Ötzi the Iceman G2a2a2 (L91) 5 kya
Notable large expansions and migrations in early history:
Indo-European migrations 6 kya from Eurasian Pontic-Caspian steppe
Astronesin migrations 5 kya from Taiwan
Bantu expansions 4 kya from Western Africa
Arab conquests 7th century from Arabian Peninsula
Below pictures show the modern descendants of haplogroups A,B,C,D,E without F which constitutes most paternal lineages outside of Africa. Some pictures were taken by me and others were downloaded from internet. The picks were based upon the ethnic aspects of the humans. Fig (1): Modern descendants of Haplogroups C,D stayed out of Africa 單倍群C,D離開非洲的現代人類後代:
Fig (2): Modern descendants of Haplogroups A,B,E stayed in Africa 單倍群A,B,E留在非洲的現代人類後代:
吳偉榮2019夏 - revived 2022冬春
Nov 12, 2021
現代東亞人的起源與形成 Origin and formation of modern East Asians
最近的基因研究表明,現代人類在東亞的人口, 比來自南方的單一起源更為複雜,通過北方路線的遺傳貢獻可能相當大。人類白細胞抗原基因的計算模擬, 支持東亞人口定居的兩種主要途徑。
Recent genetic studies have suggested that the population of East Asia by modern humans was more complex than a single origin from the South, and that a genetic contribution via a Northern route was probably quite substantial. Computer simulation of human leukocyte antigen genes supports two main routes of colonization by human populations in East Asia.
Population genomic studies have studied into the origin and formation of modern East Asians. Ancestors of East Asians split from other human populations between 70,000 to 50,000 years ago. Possible routes into East Asia by the ancestors of East Asian-related populations include a northern route model from Central Asia, and a southern route model from Southeast Asia. A third route into Northern Siberia was used by a distinct population, represented by the Ancient North Eurasians (古北歐亞人, see below post), which however did not contribute ancestry to modern East Asians. These Ancient North Eurasians however received significant geneflow of an early East Asian-related population, contributing about 22% ancestry towards the ANE, samplified by Mal'ta–Buret' 1 sample.


Fig. (1): Possible route(s) of modern human migrations towards East Asia according to different hypotheses proposed by geneticists (“pincer” or “overlapping” model: both the northern and the southern routes; “southern origin” model: only the southern route), along with representative archaeological sites during the critical period (100,000– 20,000 BP), knowing that the shallow parts of the sea (light gray/blue on the map) were postulated as land area with the lower sea level of last ice age (Sun et al. 2000). References for the archaeological sites: Mal’ta: Vasil’ev (1993); Upper Cave: Chen et al. (1989); Tianyuan Cave: Shang et al. (2007); Huanglong Cave: Wu et al. (2006); Liujiang: Shen et al. (2002); Callao Cave: Mijares et al. (2010).
Figure (2): 人類白細胞抗原基因的計算 Computer simulation of human leukocyte antigen genes supports two main routes of colonization by human populations in East Asia (2015): Topographic map of East Asia showing hypothesized modern human migration routes and suggested barriers. Triangles indicate representative Upper Paleolithic archeological sites with human remains: a. Mal’ta; b. Afontova Gora-2; c. Upper Cave; d. Tianyuan Cave; e. Liujiang; f. Chochen; g. Minatogawa; h. Niah Cave and points indicate modern populations samples for HLA−A, −B, −DRB1 loci: 1. Tuvinians; 2. Oold; 3. Khalkha; 4. Mongolians; 5. Liaoning Han; 6. Shanxi Han; 7. Shandong Han; 8. Xi’an Han; 9. Henan Han; 10. Anhui Han; 11. Hubei Han; 12. Zhejiang Han; 13. Hunan Han; 14. Jiangxi Han; 15. Fujian Han; 16. Maonan; 17. Guangdong Han; 18. Zhuang; 19. Muong.
Modern Northeast Asians consist mostly of the "Northern East Asian" component which has a continuity in Northeast Asia since at least 44,000 BC, and expanded massively with millet cultivation. Modern Southeast Asians consist mostly of the "Southern East Asian" component, which is associated with the spread of rice cultivation. Modern East Asians are positioned in between these two distinct, but closely related lineages.
Figure (3): Proposed migration routes of East-Eurasian paternal haplogroups (C, D, N, O, and Q), during the peopling of East Asia according to the "Northern Route Model". For "Southern Route Model" see earlier post: 早期人類沿海遷徙 Early Human Coastal Migrations.

Population genomic studies have studied into the origin and formation of modern East Asians. Ancestors of East Asians split from other human populations between 70,000 to 50,000 years ago. Possible routes into East Asia by the ancestors of East Asian-related populations include a northern route model from Central Asia, and a southern route model from Southeast Asia. A third route into Northern Siberia was used by a distinct population, represented by the Ancient North Eurasians (古北歐亞人, see below post), which however did not contribute ancestry to modern East Asians. These Ancient North Eurasians however received significant geneflow of an early East Asian-related population, contributing about 22% ancestry towards the ANE, samplified by Mal'ta–Buret' 1 sample.
Fig. (1): Possible route(s) of modern human migrations towards East Asia according to different hypotheses proposed by geneticists (“pincer” or “overlapping” model: both the northern and the southern routes; “southern origin” model: only the southern route), along with representative archaeological sites during the critical period (100,000– 20,000 BP), knowing that the shallow parts of the sea (light gray/blue on the map) were postulated as land area with the lower sea level of last ice age (Sun et al. 2000). References for the archaeological sites: Mal’ta: Vasil’ev (1993); Upper Cave: Chen et al. (1989); Tianyuan Cave: Shang et al. (2007); Huanglong Cave: Wu et al. (2006); Liujiang: Shen et al. (2002); Callao Cave: Mijares et al. (2010).
Figure (2): 人類白細胞抗原基因的計算 Computer simulation of human leukocyte antigen genes supports two main routes of colonization by human populations in East Asia (2015): Topographic map of East Asia showing hypothesized modern human migration routes and suggested barriers. Triangles indicate representative Upper Paleolithic archeological sites with human remains: a. Mal’ta; b. Afontova Gora-2; c. Upper Cave; d. Tianyuan Cave; e. Liujiang; f. Chochen; g. Minatogawa; h. Niah Cave and points indicate modern populations samples for HLA−A, −B, −DRB1 loci: 1. Tuvinians; 2. Oold; 3. Khalkha; 4. Mongolians; 5. Liaoning Han; 6. Shanxi Han; 7. Shandong Han; 8. Xi’an Han; 9. Henan Han; 10. Anhui Han; 11. Hubei Han; 12. Zhejiang Han; 13. Hunan Han; 14. Jiangxi Han; 15. Fujian Han; 16. Maonan; 17. Guangdong Han; 18. Zhuang; 19. Muong.
Modern Northeast Asians consist mostly of the "Northern East Asian" component which has a continuity in Northeast Asia since at least 44,000 BC, and expanded massively with millet cultivation. Modern Southeast Asians consist mostly of the "Southern East Asian" component, which is associated with the spread of rice cultivation. Modern East Asians are positioned in between these two distinct, but closely related lineages.
Figure (3): Proposed migration routes of East-Eurasian paternal haplogroups (C, D, N, O, and Q), during the peopling of East Asia according to the "Northern Route Model". For "Southern Route Model" see earlier post: 早期人類沿海遷徙 Early Human Coastal Migrations.
Jun 6, 2021
單倍羣 Haplogroup CT 2022-2-14
Updated 2022-2-14 from Wikipedia.
- Haplogroup CT (M168/PF1416)
- CF (P143/PF2587, CTS3818/M3690/PF2668, CTS6376/M3711/PF2697) Shan people
- C (M130/Page51/RPS4Y711, M216) Asia, Oceania, and North America
- C1 F3393/Z1426
- C1a CTS11043
- C1a1 formerly C1 (M8, M105, M131) Japan
- C1a2 Very low frequency in Western Eurasia (including Kostenki 38 000 years ago) and North Africa
- C1b
- C1b1
- C1b1a
- C1b1a1 formerly C5 (M356) South Asia, Central Asia, and Southwest Asia
- C1b1a
- C1b2
- C1b2a formerly C2 (M38) Indonesia, New Guinea, Melanesia, Micronesia, and Polynesia
- C1b2b formerly C4 (M347) Indigenous Australians
- C1b1
- C1a CTS11043
- C2 formerly C3 (M217, P44) Found throughout Eurasia and North America, but especially among Mongols, Kazakhs, Tungusic peoples, Paleosiberians, and Na-Dené-speaking peoples
- C1 F3393/Z1426
- F (M89, M213) Found throughout Eurasia, Oceania, and the Americas
- F1 (P91, P104)
- F2 (M427, M428)
- F3 (P96)
- F4 (M481)
- GHIJK F1329/M3658/PF2622/YSC0001299, CTS2254/M3680/PF2657, FGC2045/Z12203 Throughout Eurasia; also The Americas; at low levels/among minorities in Africa and Oceania
- G M201, P257 Primarily the Caucasus; also at low levels/among minorities in Europe, North Africa, South West Asia and Central Asia
- HIJK Throughout Eurasia; also The Americas; at low levels/among minorities in Africa and Oceania
- H M69, M370 South Asia, Central Asia, and Southwest Asia
- IJK L15/S137, L16/S138 Eurasia, North Africa, Oceania, the Americas and East Africa
- IJ M429/P125 Europe, Western Asia, North Africa and East Africa
- I M170, M258, P19, P38, P212, U179 Europe
- J 12f2.1, M304 Europe, Western Asia, South Asia, North Africa and East Africa
- K M9 Found all over Eurasia; also significant in the Americas and Oceania; at lower levels in North Africa and East Africa
- LT (K1) L298/P326
- L M11, M20, M22, M61, M185, M295 the Indian subcontinent
- T M70, M184/USP9Y+3178, M193, M272 (formerly K2) Southwestern Asia, South Asia, North Africa, the Horn of Africa, the Chad Basin, and Southern Europe
- K2 M526 Eurasia, Oceania, The Americas, some indigenous Australians and some minorities in Africa
- NO (K2a) M214 Reportedly found in small numbers of Buyi and Japanese males.
- N M231 Eastern Europe, North Asia, and East Asia
- O M175 Oceania, Southeast Asia, and East Asia
- K2b formerly MPS P331 Eurasia, The Americas, Oceania and minorities in Africa
- K2b1 formerly MS P397/P399 Melanesians, Micronesians, indigenous Australians and Polynesians.
- M P256 Eastern Indonesia, Papua New Guinea, and Melanesia
- S M230, P202, P204 (formerly K5) Eastern Indonesia, Papua New Guinea, and Melanesia
- P (K2b2) 92R7, M45, M74/N12, P27.1/P207 Throughout Eurasia, The Americas and some minorities in Africa
- K2b1 formerly MS P397/P399 Melanesians, Micronesians, indigenous Australians and Polynesians.
- K2c P261 Low levels in Bali
- K2d P402/P403 Low levels in Java
- K2e M147 Low levels in South Asia
- NO (K2a) M214 Reportedly found in small numbers of Buyi and Japanese males.
- LT (K1) L298/P326
- C (M130/Page51/RPS4Y711, M216) Asia, Oceania, and North America
- Haplogroup DE (M1/YAP, M145/P205, M203/Page36, P144, P153, P165, P167, P183) Asia, Africa, Southern and Eastern Europe; also at low levels in Oceania
- Haplogroup D (M174) Primarily Japan, Tibet and Andaman Islanders; also at low level/among minorities in East Asia, Central Asia, Micronesia and Melanesia
- Haplogroup D1 (CTS11577) Mainly Tibet; minorities in Central Asia, East Asia, and South East Asia
- Haplogroup D1a Z27276 East and Central Asia
- D1a1 (M15) (ex-D1) Qiang people; also at low levels throughout East and Central Asia
- Haplogroup D1a2 P99 (ex-D3) Tibet and Central Asia
- Haplogroup D1b (ex-D2) (M64.1/Page44.1, M55, M57, M179/Page31, M359.1/P41.1, P37.1, P190, 12f2.2) Mainly Japan; also at lowel levels/among minorities in Korea, China, Micronesia and Melanesia
- Haplogroup D1a Z27276 East and Central Asia
- Haplogroup D2 L1366 (ex-D1a) Philippines
- Haplogroup D1 (CTS11577) Mainly Tibet; minorities in Central Asia, East Asia, and South East Asia
- Haplogroup E (M40, M96) Africa, Middle East, Southern and Eastern Europe
- Haplogroup E1 (P147) At high levels throughout Africa; at lower levels in the Middle East and Europe
- Haplogroup E1a (formerly E1) (M132) Primarily Africa
- Haplogroup E1b (P177/PF1939)
- Haplogroup E1b1 (formerly E3) (P2, DYS391p)
- Haplogroup E1b1a (V38)
- Haplogroup E1b1a1 (formerly E3a) (M2) West Africa, Central Africa, Southeast Africa and Southern Africa
- Haplogroup E1b1b (formerly E3b) (M215) Horn of Africa (Ethiopians, Somalis), North Africa (Berbers, Arabs[9]), the Middle East, Europe (esp. areas near the Mediterranean Sea)
- Haplogroup E1b1 (formerly E3) (P2, DYS391p)
- Haplogroup E2 (M75) East Africa
- Haplogroup E1 (P147) At high levels throughout Africa; at lower levels in the Middle East and Europe
- Haplogroup D (M174) Primarily Japan, Tibet and Andaman Islanders; also at low level/among minorities in East Asia, Central Asia, Micronesia and Melanesia
May 20, 2017
基因檢測: 祖源分析,製造"太陽的後裔"? DNA: Ancestry,Descendants of the sun
近來有些朋友做了基因檢測,對檢測報告提出了疑問。例如有人的祖源有25%的泰國人,或30%的韓國人,但他們認為他們的上幾代從來沒有到過泰國或韓國,這是怎麼回事呢? 也有人測不出是漢人的! 我的從前基因報告,主要是談Y染色體父系祖先, 但這只是一個祖先,你還有2的N次方個祖先, 所以不一定與人種或血緣有關係。 人體的遺傳信息多數在常染色體和X染色體上, 只不過在下一代它們會發生重組(Recombination), 沒法用來追踪早期祖先的來源。
每個人體細胞核內有23對染色體,其中22對常染色體,一對性染色體(XX或XY), 此外細胞裏還有一種較小的環狀mtDNA。全組約有30億個DNA鹼基對, 當鹼基對發生變異(約佔0.5%), 從而決定下一代的遺傳。 基因公司通過基因測序(Gene Sequencing)和基因芯片篩查(Gene Chip Screening), 幫助用戶了解與基因相關的遺傳特徵,祖源和親緣關係,以及一些遺傳性疾病,健康和藥物反應。 FDA前一段時間,停止了23andMe公司提供健康方面服務, 但現在已經恢復了。 健康風險分析是未來趨勢, 這裡我只談祖源分析。
因爲只有男性有Y染色體,因此只能從他的父親傳遞下來。而mtDNA,不論男女都是繼承自母親, 它們可以用來探索族群起源與移徙。 復旦大學在2001年發了一篇文章,在整個東亞、西伯利亞、東南亞、太平洋島嶼, 測了12000個男姓的Y染色體,發現都屬於走出非洲的支系(突變標記M168:一路發)。 直到今天,東亞也沒找到一個不是走出非洲的樣本, 全世界不包括非洲裔也沒有。
歐亞現代人基因有尼安德特人和丹尼索瓦古人類混進來的, 這是否推翻了走出非洲的理論?可以說沒有。 這種混合充其量4%, 而絕大部分歐亞現代人基因仍然是和非洲各種現代人屬於同一支的, 混合程度遠達不到“多地區起源連續進化附帶雜交”的程度。另外,從考古學所見的200萬年內各大洲的古人類骨骼特徵的多態性來說,隨著時閒的發展,多態性也是明顯縮小的, 而如果是近來獨立發展爲主的話,各大洲之閒人類的分化應該越來越顯著才對。
 至於常染色體,在減數分裂的時候,是把父母兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。 有性生殖(減數分裂)重組, 使後代基因多樣化, 它發生距今約12億年, 在真核生物(人類也屬此)進化上, 現在依然持續存在。
至於常染色體,在減數分裂的時候,是把父母兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。 有性生殖(減數分裂)重組, 使後代基因多樣化, 它發生距今約12億年, 在真核生物(人類也屬此)進化上, 現在依然持續存在。
左圖例子顯示了兩個表姐弟的一對常染色體,有一共祖片段(IBD Segment), 是來自一個祖父。 一般情況下, 所有的22對常染色體, 有來自祖父或祖母不同樣的共祖片段, 但也可能沒有。 所以從理論上講,即使是親生兄弟姐妹也可能沒有任何相關性。
推算兩人的關係, 首先找到兩人的共祖片段,然後根據共祖片段的長度和數量,來判斷兩個人在幾代之內的親疏關係。 雖然判斷結果在大多數情況下是一致的,但有時有人會存在著很大的差異, 基因公司會用各種先進的技術和分析來校驗。
祖源分析其實是一個常染色體比較的過程,將你的DNA數據和不同人群的進行比較, 來看你和那些人群最相近、你可以模擬成由那些人群, 按某種祖源成分(Ancestry Composition)混合而成。
因為要比較,所以參考數據庫至關重要。 可能是由於缺少參考數據,23andme在東亞的解析度很低, 只能將你劃分為一類華人。 目前有中國基因公司分很細: 南北方漢族、納西/彝族、苗瑤語族群、藏族、拉祜族、蒙古語族群、維吾爾族、傣族、通古斯族群、畬族、高山族群, 不知道多準。 早期祖源的估計是非常不準確的,但已經大大改善了。 通常大地區的結果是準確的,但更具體的分析可能不準, 例如歐洲人往往會誇大Scandinavian的比例。
上面提到的泰國人祖源,是參考人群細分問題。 壯、泰、老、傣、掸族等同屬一個語系, 國際上叫 Tai–Kadai languages, 中文叫侗台或壯侗語系, 是被認為來自古代百越的語言。 吳越地(江浙)被認為有1/4百越血統, 上述泰國人祖源可能來自百越。 以前我提到古代越人擁楫歌, 原文以漢字記音, 被現代侗台語(泰國語)破譯, 證明侗台語來自吳越。
上面提到的韓國人祖源,是參考數據辨析問題。 如果只有現代人群數據,那麼很難搞清楚誰是“源”誰是“流”,引入古代人群作為參考數據,可以在一定程度上幫助解決基因流向問題。 除非我們可以收集到很多很多從古到今, 各個時間點的大量漢人和朝鮮人的DNA樣本,才能確定數據有多大機率來自朝鮮人或漢人祖先。 如今世界上一些少數部落民族人口逐年減少, 他們純正的基因數據,對基因流向非常重要。
根據某基因檢測公司首席科學家, 韓國人的基因可按照漢族:烏爾奇=7:3這個比率。 如果有韓劇女粉絲, 想要製造出一個"太陽的後裔"宋仲基, 可以這樣做。 比如妳是漢人,那麼妳要先到俄羅斯遠東地區, 找到一個烏爾奇人(Ulchi, 是純正的通古斯族),嫁他生孩子,然後讓妳的孩子再嫁或娶一個漢人,這樣妳的孫子就和宋仲基很相似了。 如今烏爾奇人數不多了, 請快! 不好意思,我沒看過這連續劇。

為什麼有人測不出是漢人? 首先要知道什麼是漢人。 我認為漢人是近代的觀念, 是社會認可的一人群, 如果社會說你是漢人,那麼你就是漢人。 如果用語言和文化來分,這是不正確的,因為一些漢人不會說漢語或有漢文化。 在歷史上漢化少數民族,現在是漢人。
我的其他輔助資料鏈接:
父系祖先, 請看 人類的單倍羣、遷徙、歷史
東亞族群, 請看 東亞民族單倍羣分佈
越人擁楫歌破譯: 吳語的源流
注: Y-單倍羣分類的精度每年都在細化, 2012年發現了F75突變位點, 是O1和O2的共同祖先。那麼按照命名原則,原來的O1和O2成爲新的O1a和O1b,而原先的O3改爲O2。為方便起見,我們經常使用原版本,也就是O1-M119,O2-M268,O3-M122。只需要註明所用的定義位點,比如說O-M122,這樣就不會混了。

吳偉榮2017夏
每個人體細胞核內有23對染色體,其中22對常染色體,一對性染色體(XX或XY), 此外細胞裏還有一種較小的環狀mtDNA。全組約有30億個DNA鹼基對, 當鹼基對發生變異(約佔0.5%), 從而決定下一代的遺傳。 基因公司通過基因測序(Gene Sequencing)和基因芯片篩查(Gene Chip Screening), 幫助用戶了解與基因相關的遺傳特徵,祖源和親緣關係,以及一些遺傳性疾病,健康和藥物反應。 FDA前一段時間,停止了23andMe公司提供健康方面服務, 但現在已經恢復了。 健康風險分析是未來趨勢, 這裡我只談祖源分析。
因爲只有男性有Y染色體,因此只能從他的父親傳遞下來。而mtDNA,不論男女都是繼承自母親, 它們可以用來探索族群起源與移徙。 復旦大學在2001年發了一篇文章,在整個東亞、西伯利亞、東南亞、太平洋島嶼, 測了12000個男姓的Y染色體,發現都屬於走出非洲的支系(突變標記M168:一路發)。 直到今天,東亞也沒找到一個不是走出非洲的樣本, 全世界不包括非洲裔也沒有。
歐亞現代人基因有尼安德特人和丹尼索瓦古人類混進來的, 這是否推翻了走出非洲的理論?可以說沒有。 這種混合充其量4%, 而絕大部分歐亞現代人基因仍然是和非洲各種現代人屬於同一支的, 混合程度遠達不到“多地區起源連續進化附帶雜交”的程度。另外,從考古學所見的200萬年內各大洲的古人類骨骼特徵的多態性來說,隨著時閒的發展,多態性也是明顯縮小的, 而如果是近來獨立發展爲主的話,各大洲之閒人類的分化應該越來越顯著才對。
 至於常染色體,在減數分裂的時候,是把父母兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。 有性生殖(減數分裂)重組, 使後代基因多樣化, 它發生距今約12億年, 在真核生物(人類也屬此)進化上, 現在依然持續存在。
至於常染色體,在減數分裂的時候,是把父母兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。 有性生殖(減數分裂)重組, 使後代基因多樣化, 它發生距今約12億年, 在真核生物(人類也屬此)進化上, 現在依然持續存在。
左圖例子顯示了兩個表姐弟的一對常染色體,有一共祖片段(IBD Segment), 是來自一個祖父。 一般情況下, 所有的22對常染色體, 有來自祖父或祖母不同樣的共祖片段, 但也可能沒有。 所以從理論上講,即使是親生兄弟姐妹也可能沒有任何相關性。
推算兩人的關係, 首先找到兩人的共祖片段,然後根據共祖片段的長度和數量,來判斷兩個人在幾代之內的親疏關係。 雖然判斷結果在大多數情況下是一致的,但有時有人會存在著很大的差異, 基因公司會用各種先進的技術和分析來校驗。
祖源分析其實是一個常染色體比較的過程,將你的DNA數據和不同人群的進行比較, 來看你和那些人群最相近、你可以模擬成由那些人群, 按某種祖源成分(Ancestry Composition)混合而成。
因為要比較,所以參考數據庫至關重要。 可能是由於缺少參考數據,23andme在東亞的解析度很低, 只能將你劃分為一類華人。 目前有中國基因公司分很細: 南北方漢族、納西/彝族、苗瑤語族群、藏族、拉祜族、蒙古語族群、維吾爾族、傣族、通古斯族群、畬族、高山族群, 不知道多準。 早期祖源的估計是非常不準確的,但已經大大改善了。 通常大地區的結果是準確的,但更具體的分析可能不準, 例如歐洲人往往會誇大Scandinavian的比例。
上面提到的泰國人祖源,是參考人群細分問題。 壯、泰、老、傣、掸族等同屬一個語系, 國際上叫 Tai–Kadai languages, 中文叫侗台或壯侗語系, 是被認為來自古代百越的語言。 吳越地(江浙)被認為有1/4百越血統, 上述泰國人祖源可能來自百越。 以前我提到古代越人擁楫歌, 原文以漢字記音, 被現代侗台語(泰國語)破譯, 證明侗台語來自吳越。
上面提到的韓國人祖源,是參考數據辨析問題。 如果只有現代人群數據,那麼很難搞清楚誰是“源”誰是“流”,引入古代人群作為參考數據,可以在一定程度上幫助解決基因流向問題。 除非我們可以收集到很多很多從古到今, 各個時間點的大量漢人和朝鮮人的DNA樣本,才能確定數據有多大機率來自朝鮮人或漢人祖先。 如今世界上一些少數部落民族人口逐年減少, 他們純正的基因數據,對基因流向非常重要。
根據某基因檢測公司首席科學家, 韓國人的基因可按照漢族:烏爾奇=7:3這個比率。 如果有韓劇女粉絲, 想要製造出一個"太陽的後裔"宋仲基, 可以這樣做。 比如妳是漢人,那麼妳要先到俄羅斯遠東地區, 找到一個烏爾奇人(Ulchi, 是純正的通古斯族),嫁他生孩子,然後讓妳的孩子再嫁或娶一個漢人,這樣妳的孫子就和宋仲基很相似了。 如今烏爾奇人數不多了, 請快! 不好意思,我沒看過這連續劇。

為什麼有人測不出是漢人? 首先要知道什麼是漢人。 我認為漢人是近代的觀念, 是社會認可的一人群, 如果社會說你是漢人,那麼你就是漢人。 如果用語言和文化來分,這是不正確的,因為一些漢人不會說漢語或有漢文化。 在歷史上漢化少數民族,現在是漢人。
我的其他輔助資料鏈接:
父系祖先, 請看 人類的單倍羣、遷徙、歷史
東亞族群, 請看 東亞民族單倍羣分佈
越人擁楫歌破譯: 吳語的源流
注: Y-單倍羣分類的精度每年都在細化, 2012年發現了F75突變位點, 是O1和O2的共同祖先。那麼按照命名原則,原來的O1和O2成爲新的O1a和O1b,而原先的O3改爲O2。為方便起見,我們經常使用原版本,也就是O1-M119,O2-M268,O3-M122。只需要註明所用的定義位點,比如說O-M122,這樣就不會混了。

吳偉榮2017夏
Sep 26, 2016
各方言區漢族的父系祖先 Paternal Ancestors of Chinese of Different Dialects
上次我們講大約6千年前,有3個人是現在四成多華裔的祖先(之前Post),主要是說漢族,但不一定是漢族。 這三個人是實際存在的,被稱爲“農民甲”、“農民乙”和“農民丙”, 或者稱爲“泰皇”、“地皇” 和“天皇”。 短時閒也不容易說明他們是什麼人,或居住什麼地方, 那麼誰是他們的後代呢?
上次也說了許多族羣定義是在語言, 例如漢語下有各種方言族羣。 如果不按行政單位劃分,而按語言區劃分(下面附錄圖片),方言之閒的Y染色體, 是否會出現分佈上的不同? 根據復旦大學圖片資料(下面附錄圖表),我計量了漢族按方言區的Y單倍羣分佈(有關Haplogroup看:之前Post), 列表如下(單位是%):

(資料僅供參考, 不宜過度解讀)
由上表顯示各方言區, 3支後代是有不同分佈的,除了中原官話區稍高, 但總計(主要為農民甲、乙、丙)約40%左右, 它們的意義仍在調查中。 同時閩語區的農民甲支系的數據偏高29.6%, 可能與下面相關: 復旦大學測到的福建和海南姓林的漢族, 有超過90%都是農民甲後代,而不姓林的漢族只有17%,而其他地方姓林的也不高, 這種姓氏和Y染色體在較大地區內, 對應得這麼好的並不多見。
上表也包括O3型(O3-M122, 上3支在內),平均約占漢族55%, 還有O型(O-M175, O=O1+O2+O3), 平均約占80%。 南方漢人O型比較高, 是來自O1和O2。 比較特別的是, 晉語區有12.5%Q型(印第安人主要類型, 下面粉紅色線條), 東北官話區有4.7%R型(印歐人主要類型, 下面粉綠色線條)。 約2萬年前,Q和R型人群在歐亞這一帶活躍,一個向東走,一個向西走。 之前我談到, 中國發現古DNA(2~3千年前),有一些貴族為Q型, 孔子也有可能是Q型(之前Post)。
閱讀我過去的文章應該知道,單倍羣和人種不一定有因果關係,但可探索族群起源與移徙,了解人類歷史。 漢族是漢朝後的名稱,華夏人名稱源於周朝,可能指當時的貴族,不一定是現代漢族的直系祖先。 近來網上有一些遺傳人類學文章報導,內容錯誤極多,希望大家不要誤讀。 Haplogroup and race do not necessarily have a causal relationship, but you can explore the human origin, migration and history.
最近我兒子曾給我發email有關中國歷史的特別文章: Chinese Civilization Come From Ancient Egypt, 意思商人來自古埃及, 我對此不置可否, 但考慮商人是什麼人種? 比如甲骨文的鬼方, 很可能是印歐人, 當時鬼字的含義,也許不是Ghost。 又如好字,不是Good意思, 是"子方之女"的含義。 子方是什麼人種? 我也曾報導, 中國古代文法比較像英文,甲骨文叫王亥(如英文的King Arthur)而不叫亥王, 又如"我祭祀於安陽"(I worship at Anyang)而不是"我在安陽祭祀"。 有興趣請看: 開始商朝 Beginning of Shang.
在閩語區, 有稱Hohlo人的語言族羣, "Hohlo"一詞的來源是什麼呢? 有興趣請看: Hohlo的來源
Apendix 附錄
下面圖片是復旦大學現代人類學, 根據漢族各方言區的2015更新數據。 方言區的分法定義如下: 其中紅色邊界是漢語十大方言邊界,藍色爲官話內部的方言區邊界。

方言定得不一定很準,比如蘇南、廣東,或者專門去的湖南等地,方言記錄應該是精確的。 北方的一些地區,方言沒有專門記,只是大致按籍貫地所屬方言算。 圖中每一行的樣本最少50個,最多幾百個, 同宗只按一個樣本統計。

吳偉榮2016秋
上次也說了許多族羣定義是在語言, 例如漢語下有各種方言族羣。 如果不按行政單位劃分,而按語言區劃分(下面附錄圖片),方言之閒的Y染色體, 是否會出現分佈上的不同? 根據復旦大學圖片資料(下面附錄圖表),我計量了漢族按方言區的Y單倍羣分佈(有關Haplogroup看:之前Post), 列表如下(單位是%):

(資料僅供參考, 不宜過度解讀)
由上表顯示各方言區, 3支後代是有不同分佈的,除了中原官話區稍高, 但總計(主要為農民甲、乙、丙)約40%左右, 它們的意義仍在調查中。 同時閩語區的農民甲支系的數據偏高29.6%, 可能與下面相關: 復旦大學測到的福建和海南姓林的漢族, 有超過90%都是農民甲後代,而不姓林的漢族只有17%,而其他地方姓林的也不高, 這種姓氏和Y染色體在較大地區內, 對應得這麼好的並不多見。
上表也包括O3型(O3-M122, 上3支在內),平均約占漢族55%, 還有O型(O-M175, O=O1+O2+O3), 平均約占80%。 南方漢人O型比較高, 是來自O1和O2。 比較特別的是, 晉語區有12.5%Q型(印第安人主要類型, 下面粉紅色線條), 東北官話區有4.7%R型(印歐人主要類型, 下面粉綠色線條)。 約2萬年前,Q和R型人群在歐亞這一帶活躍,一個向東走,一個向西走。 之前我談到, 中國發現古DNA(2~3千年前),有一些貴族為Q型, 孔子也有可能是Q型(之前Post)。
閱讀我過去的文章應該知道,單倍羣和人種不一定有因果關係,但可探索族群起源與移徙,了解人類歷史。 漢族是漢朝後的名稱,華夏人名稱源於周朝,可能指當時的貴族,不一定是現代漢族的直系祖先。 近來網上有一些遺傳人類學文章報導,內容錯誤極多,希望大家不要誤讀。 Haplogroup and race do not necessarily have a causal relationship, but you can explore the human origin, migration and history.
最近我兒子曾給我發email有關中國歷史的特別文章: Chinese Civilization Come From Ancient Egypt, 意思商人來自古埃及, 我對此不置可否, 但考慮商人是什麼人種? 比如甲骨文的鬼方, 很可能是印歐人, 當時鬼字的含義,也許不是Ghost。 又如好字,不是Good意思, 是"子方之女"的含義。 子方是什麼人種? 我也曾報導, 中國古代文法比較像英文,甲骨文叫王亥(如英文的King Arthur)而不叫亥王, 又如"我祭祀於安陽"(I worship at Anyang)而不是"我在安陽祭祀"。 有興趣請看: 開始商朝 Beginning of Shang.
在閩語區, 有稱Hohlo人的語言族羣, "Hohlo"一詞的來源是什麼呢? 有興趣請看: Hohlo的來源
Apendix 附錄
下面圖片是復旦大學現代人類學, 根據漢族各方言區的2015更新數據。 方言區的分法定義如下: 其中紅色邊界是漢語十大方言邊界,藍色爲官話內部的方言區邊界。

方言定得不一定很準,比如蘇南、廣東,或者專門去的湖南等地,方言記錄應該是精確的。 北方的一些地區,方言沒有專門記,只是大致按籍貫地所屬方言算。 圖中每一行的樣本最少50個,最多幾百個, 同宗只按一個樣本統計。

吳偉榮2016秋
Jul 30, 2016
6千年前有3人是四成多華裔的祖先 Three neolithic men are 40% Chinese ancestors
由於新一代基因測序(Sequencing)的成本顯著下降,可做深覆蓋(Coverage)的大量個體測序, 基因大數據將是未來醫療的發展方向。目前只要少於2百美元, 就可分析出個人的祖源、遺傳結構,並根據當前的數據計算出個人的健康風險、遺傳疾病、天賦特徵和藥物反應, 並構建個性化醫療(Personalized Medicine),所以對華裔基因了解是很重要的。
大約6千年前左右, 有3個人是現在四成多華裔的祖先, 這裡主要是說漢族, 也包括一些少數民族和鄰近國家民族。我們檢測的是Y染色體父系祖先, 因爲Y染色體位點多,平均每100年就能有一個突變,而母系用線粒體平均要2000多年, 才能有一次突變。 當然不是說他們的族羣沒有其他男性,而是說這些男性都沒能直接傳下男性後代,只有這三個幸運兒的後代(包括女性), 終於活到了現代。
到底這三個祖先是什麼人? 首先這個擴張年代正好在新石器時代晚期, 這個時候穀類從採集、狩獵的輔助食物變成了主食,人類的食物開始依賴農業,同時聚落的分佈密度和人口大大增加, 社會從母系變成了父系社會。因此對於這三個祖先的解釋: 他們是最早的農民,因爲穀物和人口的生產力大幅發展,成了超級祖先,因此稱他們爲“農民甲”、“農民乙”和“農民丙”。
有人不相信這種解釋,認爲他們肯定是很偉大的部落首領,有很大的權力而能占有很多女人,建議用三皇來稱呼,按照計出現的時閒, 分別稱爲“泰皇”、“地皇” 和“天皇”。 還有人稱他們作“炎帝”、“黃帝”和“蚩尤”,事實上不恰當的, 他們只是神話人物,名字在戰國之後才出現。 例如說成吉思汗有很多後代, 是不靠譜的, 因爲他的遺骸古DNA從未找到。 上述三個人是客觀存在的,有科學依據的,短時閒也不容易說明他們是什麼樣的人物。 這也說明了, 漢族起源於幾千年前一個祖先是不可能的, 下面說明最大的簇, 也不過占漢族父系的16%。
Click to see larger figure:
 近來線粒體基因組的研究, 顯示東亞和歐洲的母系的人口擴張, 開始於新石器時代之前,這就與農業發展是驅動力說法矛盾。 有人認爲父系的擴張,不只與農業發展相關,也有可能與外來的小麥、綿羊、青銅武器和工具、馬的馴化及戰車的輸入相關。
近來線粒體基因組的研究, 顯示東亞和歐洲的母系的人口擴張, 開始於新石器時代之前,這就與農業發展是驅動力說法矛盾。 有人認爲父系的擴張,不只與農業發展相關,也有可能與外來的小麥、綿羊、青銅武器和工具、馬的馴化及戰車的輸入相關。
依照現有的Y染色體進化樹(左圖),非洲之外幾乎都是M168(一路發走出非洲)突變型的後代。 東亞常見的祖系, 都是在2萬年前以前分支出來的,而現已知的大部分支系, 是在舊石器時代晚期(1萬年前)產生的。 進化樹上7千年以前的分支都是二叉的,這表明在舊石器時代的人口增長緩慢, 淘汰了大部分曾經存在過的支系。
O3型約發生在2萬年前,即末次冰盛期之後的舊石器時代。 這段時閒全球氣候逐漸回暖,海平面從現在海面以下百米一直升到和目前海平面接近的高度,人類的可活動範圍大大擴展,技術上也有了一些創新,農業也出現了。
約在6千年前,O3出現了三個星簇(star cluster)的擴張,分別命名爲Oα, Oβ和Oγ單倍羣。 這三個人出現的年代很接近, 即突然從一個人演化出難以分出先後的5–7個支系,而且這些支系都是有後代一直延續到現在, 他們的後代約爲16%, 11%和14%, 總共41%。
其實許多族羣定義是在語言, 例如漢藏語系, 漢語下包含各種方言族羣。 有主張說某個方言的族羣是最純正的漢人,有說南方漢人主要是土著漢化的,有說北方人是胡種的。其實這些說法都是片面的,有很多是出於顯示本族羣的正統性, 而貶抑其他族羣而製造出來的。 本文從科學依據、數據中發現, 南北漢人的父系的結構還是接近的,如果把其他民族擺一起就能看出明顯的區別。
Appendix 附錄
(下面圖1-3)是三個包括Oα、Oβ和Oγ在內的單倍羣分佈圖(Haplogroup): O3a2c1a-M117、O3a2c1*-M134和O3a1c-002611, 這些圖均爲相互無關男性數據,以深淺標明各單倍羣所占比例。省份區域爲漢族,樣本數較少的省份作了合併, 圓圈爲注明的少數民族。 O3a2c1a-M117在藏緬族群中也有較多分佈, O3下的另一支系O3a2b-M7在苗瑤和孟高棉人群中高頻出現,但在漢族中卻不足5%。
還有非O3祖先(下面圖5和6): 一支是O1a的一個下游擴張, 在良渚文化的古DNA有很高比例,年代可能只有4000年不到。 另一支是C3的一個下游擴張(南支),擴張時閒可能比O3的三大簇略早。 蒙古人是C3北支,漢人不常見。 這兩支的人口大概也各占了漢族的10%上下,但因爲沒有足夠的樣本或測序質量不好,尚未能表現出星簇擴張。 如果把這兩支也加上,這五個新石器祖先的後代, 就能占到漢族60%的比例了。
還有一單倍羣O2a(下面圖4)出現在華南、南亞和東南亞一帶,曹操證明是這型的, 因爲找到他叔祖的古DNA。另外一分支O2b出現在日本、朝鮮和中國東北, 漢族很少。 如果包括所有的O單倍群,中國人及朝鮮人超過75%, 日本人超過50%。 日本另有高頻35%的D2型, 在朝鮮半島只有低於1%的比例, 目前在大量的漢族數據中沒測到一例。






Most materials from polyhedron 復旦大學現代人類學實驗室
(請看之前單倍群報告, Click "Home" at bottom)
吳偉榮2016夏
大約6千年前左右, 有3個人是現在四成多華裔的祖先, 這裡主要是說漢族, 也包括一些少數民族和鄰近國家民族。我們檢測的是Y染色體父系祖先, 因爲Y染色體位點多,平均每100年就能有一個突變,而母系用線粒體平均要2000多年, 才能有一次突變。 當然不是說他們的族羣沒有其他男性,而是說這些男性都沒能直接傳下男性後代,只有這三個幸運兒的後代(包括女性), 終於活到了現代。
到底這三個祖先是什麼人? 首先這個擴張年代正好在新石器時代晚期, 這個時候穀類從採集、狩獵的輔助食物變成了主食,人類的食物開始依賴農業,同時聚落的分佈密度和人口大大增加, 社會從母系變成了父系社會。因此對於這三個祖先的解釋: 他們是最早的農民,因爲穀物和人口的生產力大幅發展,成了超級祖先,因此稱他們爲“農民甲”、“農民乙”和“農民丙”。
有人不相信這種解釋,認爲他們肯定是很偉大的部落首領,有很大的權力而能占有很多女人,建議用三皇來稱呼,按照計出現的時閒, 分別稱爲“泰皇”、“地皇” 和“天皇”。 還有人稱他們作“炎帝”、“黃帝”和“蚩尤”,事實上不恰當的, 他們只是神話人物,名字在戰國之後才出現。 例如說成吉思汗有很多後代, 是不靠譜的, 因爲他的遺骸古DNA從未找到。 上述三個人是客觀存在的,有科學依據的,短時閒也不容易說明他們是什麼樣的人物。 這也說明了, 漢族起源於幾千年前一個祖先是不可能的, 下面說明最大的簇, 也不過占漢族父系的16%。
Click to see larger figure:
 近來線粒體基因組的研究, 顯示東亞和歐洲的母系的人口擴張, 開始於新石器時代之前,這就與農業發展是驅動力說法矛盾。 有人認爲父系的擴張,不只與農業發展相關,也有可能與外來的小麥、綿羊、青銅武器和工具、馬的馴化及戰車的輸入相關。
近來線粒體基因組的研究, 顯示東亞和歐洲的母系的人口擴張, 開始於新石器時代之前,這就與農業發展是驅動力說法矛盾。 有人認爲父系的擴張,不只與農業發展相關,也有可能與外來的小麥、綿羊、青銅武器和工具、馬的馴化及戰車的輸入相關。
依照現有的Y染色體進化樹(左圖),非洲之外幾乎都是M168(一路發走出非洲)突變型的後代。 東亞常見的祖系, 都是在2萬年前以前分支出來的,而現已知的大部分支系, 是在舊石器時代晚期(1萬年前)產生的。 進化樹上7千年以前的分支都是二叉的,這表明在舊石器時代的人口增長緩慢, 淘汰了大部分曾經存在過的支系。
O3型約發生在2萬年前,即末次冰盛期之後的舊石器時代。 這段時閒全球氣候逐漸回暖,海平面從現在海面以下百米一直升到和目前海平面接近的高度,人類的可活動範圍大大擴展,技術上也有了一些創新,農業也出現了。
約在6千年前,O3出現了三個星簇(star cluster)的擴張,分別命名爲Oα, Oβ和Oγ單倍羣。 這三個人出現的年代很接近, 即突然從一個人演化出難以分出先後的5–7個支系,而且這些支系都是有後代一直延續到現在, 他們的後代約爲16%, 11%和14%, 總共41%。
其實許多族羣定義是在語言, 例如漢藏語系, 漢語下包含各種方言族羣。 有主張說某個方言的族羣是最純正的漢人,有說南方漢人主要是土著漢化的,有說北方人是胡種的。其實這些說法都是片面的,有很多是出於顯示本族羣的正統性, 而貶抑其他族羣而製造出來的。 本文從科學依據、數據中發現, 南北漢人的父系的結構還是接近的,如果把其他民族擺一起就能看出明顯的區別。
Appendix 附錄
(下面圖1-3)是三個包括Oα、Oβ和Oγ在內的單倍羣分佈圖(Haplogroup): O3a2c1a-M117、O3a2c1*-M134和O3a1c-002611, 這些圖均爲相互無關男性數據,以深淺標明各單倍羣所占比例。省份區域爲漢族,樣本數較少的省份作了合併, 圓圈爲注明的少數民族。 O3a2c1a-M117在藏緬族群中也有較多分佈, O3下的另一支系O3a2b-M7在苗瑤和孟高棉人群中高頻出現,但在漢族中卻不足5%。
還有非O3祖先(下面圖5和6): 一支是O1a的一個下游擴張, 在良渚文化的古DNA有很高比例,年代可能只有4000年不到。 另一支是C3的一個下游擴張(南支),擴張時閒可能比O3的三大簇略早。 蒙古人是C3北支,漢人不常見。 這兩支的人口大概也各占了漢族的10%上下,但因爲沒有足夠的樣本或測序質量不好,尚未能表現出星簇擴張。 如果把這兩支也加上,這五個新石器祖先的後代, 就能占到漢族60%的比例了。
還有一單倍羣O2a(下面圖4)出現在華南、南亞和東南亞一帶,曹操證明是這型的, 因爲找到他叔祖的古DNA。另外一分支O2b出現在日本、朝鮮和中國東北, 漢族很少。 如果包括所有的O單倍群,中國人及朝鮮人超過75%, 日本人超過50%。 日本另有高頻35%的D2型, 在朝鮮半島只有低於1%的比例, 目前在大量的漢族數據中沒測到一例。






Most materials from polyhedron 復旦大學現代人類學實驗室
(請看之前單倍群報告, Click "Home" at bottom)
吳偉榮2016夏
Mar 16, 2016
基因和基因表現 Genetics & Epigenetics
現今在東方與西方社會裏, 都流行著的許多方面健康之道, 它與遺傳基因、生活方式和環境有很大的關係,我們希望能了解它的科學依據, 可以挑選更好的養生之道。還有,如今醫生都希望病人能夠理解更多的自己的病因,可以幫助醫生的診斷和治療。
核酸(Nucleic Acids),蛋白質(Proteins), 碳水化合物(Carbohydrates)和脂質(Lipids),都是生物體的主要大分子, 核酸包括DNA和RNA。人體每個細胞核中共有23對染色體(Chromosomes),每條染色體都有一條DNA分子。其中22對稱作常染色體(Autosomal),另外女人有一對X染色體,男人則是一條X和一條Y。此外,細胞裏還有一種環狀DNA,簡寫mtDNA。
因爲只有男人有Y染色體,因此只能從他的父親傳遞下來。而mtDNA,不論男女都是繼承自母親。至於常染色體,在減數分裂的時候,是把兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。一對X染色體在減數分裂時, 同樣也會重組。因此我們無法通過常染色體或X染色體來追蹤祖先。
DNA是由四種核苷酸(Nucleotides)連接成為雙鏈螺旋鏈, 核苷酸序列(Sequence)就是生物體的遺傳資訊, 而基因(Gene)則是一組連續的核苷酸序列。每條染色體上都帶有一定數量的基因。一個基因在細胞有絲分裂時有兩個對列的位點,稱為等位基因(Alleles),分別來自父與母。依所攜帶性狀(Trait)的表現,又可分為顯性(Dominant)基因和隱性(Recessive)基因。

等位基因可在研究花色的實驗中理解,豌豆花的顏色只有兩種:紫色和白色,卻沒有任何一朵顯示出兩種顏色的中間色。每一顆豌豆都可以從父母分別繼承其中的一個等位基因, 常常有一個是顯性的,決定了生物體的表現型,而另一個就被稱為隱性的,不會被表現出來。左圖顯示了帶有兩種花色基因(顯性紫色B和隱性白色b)的豌豆雜交後的結果。
另一例子是在之前我的Post, 談到耳垢(體味)的等位基因, 在全世界各人羣的分佈比例非常不同。 因為發生基因突變,東亞人是沒味的,而新疆姑娘是半臭或半香的。如果有人想要找配偶,可以參考裡面的世界地圖。(看下面的Post)
細胞以基因的DNA序列為模板, 被轉錄為RNA副本, 以三個核苷酸為一組,用於產生特定的蛋白質的胺基酸(Amino acid)序列, 這種對應性被稱為遺傳密碼(Genetic code), 胺基酸序列決定了蛋白質的結構與它的功能。基因轉錄是可被調控的, 細胞中存在許多方式來控制基因表現(Gene expression),以便產生符合細胞需求的蛋白質。

表徵遺傳學(Epigenetics)是與遺傳學(Genetics)相對應的概念, 是1980年代逐漸興起的一門學科,在不改變DNA序列的前提下, 通過某些機制引起可遺傳的基因表現的變化。機制包括DNA甲基化(Methylation)、染色質重構、RNA調控等, 是受到飲食、情緒、化學物質等影響。如何打開優良基因,關閉不良基因, 未來表觀遺傳學將作為一個治療疾病的重要研究領域。

2008年美國NIH投資1.9億美元用於Epigenetics研究, 去了解老化機制,人類發育和癌症起源,心臟病,精神疾病及其他的健康狀況的潛力。 Time雜誌於 2010 年 發表了一個特別報導 "為何你的基因不是命運的主宰(Why Your DNA Isn’t Your Destiny)" 雖然有點過時,但它很容易閱讀。(看網頁下面最後的Post)
All inserted images are from Wikipedia.
養生之道請看: 健康之道:活過100歲
核酸(Nucleic Acids),蛋白質(Proteins), 碳水化合物(Carbohydrates)和脂質(Lipids),都是生物體的主要大分子, 核酸包括DNA和RNA。人體每個細胞核中共有23對染色體(Chromosomes),每條染色體都有一條DNA分子。其中22對稱作常染色體(Autosomal),另外女人有一對X染色體,男人則是一條X和一條Y。此外,細胞裏還有一種環狀DNA,簡寫mtDNA。
因爲只有男人有Y染色體,因此只能從他的父親傳遞下來。而mtDNA,不論男女都是繼承自母親。至於常染色體,在減數分裂的時候,是把兩條打亂後接成一條重組。因此每個人的一對染色體通常來自四個祖父母。一對X染色體在減數分裂時, 同樣也會重組。因此我們無法通過常染色體或X染色體來追蹤祖先。
DNA是由四種核苷酸(Nucleotides)連接成為雙鏈螺旋鏈, 核苷酸序列(Sequence)就是生物體的遺傳資訊, 而基因(Gene)則是一組連續的核苷酸序列。每條染色體上都帶有一定數量的基因。一個基因在細胞有絲分裂時有兩個對列的位點,稱為等位基因(Alleles),分別來自父與母。依所攜帶性狀(Trait)的表現,又可分為顯性(Dominant)基因和隱性(Recessive)基因。

等位基因可在研究花色的實驗中理解,豌豆花的顏色只有兩種:紫色和白色,卻沒有任何一朵顯示出兩種顏色的中間色。每一顆豌豆都可以從父母分別繼承其中的一個等位基因, 常常有一個是顯性的,決定了生物體的表現型,而另一個就被稱為隱性的,不會被表現出來。左圖顯示了帶有兩種花色基因(顯性紫色B和隱性白色b)的豌豆雜交後的結果。
另一例子是在之前我的Post, 談到耳垢(體味)的等位基因, 在全世界各人羣的分佈比例非常不同。 因為發生基因突變,東亞人是沒味的,而新疆姑娘是半臭或半香的。如果有人想要找配偶,可以參考裡面的世界地圖。(看下面的Post)
細胞以基因的DNA序列為模板, 被轉錄為RNA副本, 以三個核苷酸為一組,用於產生特定的蛋白質的胺基酸(Amino acid)序列, 這種對應性被稱為遺傳密碼(Genetic code), 胺基酸序列決定了蛋白質的結構與它的功能。基因轉錄是可被調控的, 細胞中存在許多方式來控制基因表現(Gene expression),以便產生符合細胞需求的蛋白質。

表徵遺傳學(Epigenetics)是與遺傳學(Genetics)相對應的概念, 是1980年代逐漸興起的一門學科,在不改變DNA序列的前提下, 通過某些機制引起可遺傳的基因表現的變化。機制包括DNA甲基化(Methylation)、染色質重構、RNA調控等, 是受到飲食、情緒、化學物質等影響。如何打開優良基因,關閉不良基因, 未來表觀遺傳學將作為一個治療疾病的重要研究領域。

2008年美國NIH投資1.9億美元用於Epigenetics研究, 去了解老化機制,人類發育和癌症起源,心臟病,精神疾病及其他的健康狀況的潛力。 Time雜誌於 2010 年 發表了一個特別報導 "為何你的基因不是命運的主宰(Why Your DNA Isn’t Your Destiny)" 雖然有點過時,但它很容易閱讀。(看網頁下面最後的Post)
All inserted images are from Wikipedia.
養生之道請看: 健康之道:活過100歲
Dec 4, 2015
常染色體遺傳: 耳垢、初乳、狐臭、狐狸精 Autosomal: earwax, colostrum & body odor etc.
雖然Y染色體可追踪祖先, 但人體絕大多數的遺傳信息都是在常染色體和X染色體上, 只不過在下一代它們會發生重組,沒法用來追踪祖先的來源。但它們能分析出有關各種人的長相、性格、能力、疾病, 如耳垢、初乳、狐臭、粉刺、曬黑、膚色、捲髮、血型、高血壓、肥胖、瞳孔顏色、高原缺氧等。
人類走出熱帶非洲,在進入亞洲溫帶的過程中,第16號常染色體的ABCC11上發生基因突變,導致東方人的溼型耳垢(earwax)變乾, 還有母親的初乳分泌減少。 初乳(colostrum)是產仔後短時閒內分泌的乳汁,蛋白質含量比常乳高,包含有豐富的免疫球蛋白、乳鐵蛋白、生長因子等,有助於嬰兒增強體質。
溼性耳垢的人容易產生狐臭, 就是腋窩下出汗,分泌一些脂肪酸類, 這些本身氣味並不大,但被一些細菌分解, 產生一些臭的有機物。對於乾耳垢的人來說,因爲腋下脂肪酸分泌少,細菌也長得慢,哪怕幾天不洗澡也不會有狐臭。
但對於溼耳垢的人來說,在熱天只需要半天狐臭就會出現。東方人通常都是晚上洗澡,正好除去一天的灰塵睡覺。而歐洲人卻一般都是早起洗澡,如果早上不洗,晚上腋窩的汗液在上午也許就能讓細菌長起來而造成狐臭。
狐臭在全世界各人羣的分佈比例非常不同, 這個突變主要分佈在華北中原地區, 因此在中國,狐臭反而成了極少數。韓國人跟中國人差不多,但日本人當中只有84%沒有狐臭。大洋洲土著只有30%沒有狐臭,白人裡面只有10%沒有狐臭, 黑人中只有0.5%沒有狐臭。印第安人也是大多沒有狐臭,說明他們是從亞洲過來的。日本人有狐臭的人集中在阿伊努人中,說明與大和民族的不同起源。
在動物中,體味有很大作用,是吸引異性的信息素, 所以說有狐臭才是人的常態。中國古詩說女子“香汗薄衫涼”,一香一臭,看似相反,其實原理完全相同。少了淡了就香,多了濃了就臭,香奈兒5號打翻了瓶子也是要熏死人的。
中國自古就有“狐狸精”的傳說,可能與狐臭有關。在古代,西方女子皮膚白皙、身材好,同時不拘禮教,招東方男性的喜歡,自然引起東方女子的妒嫉, 認為她們是狐狸變的,所以有狐臭。狐臭本來稱作“胡臭”,是區別胡漢的一種標誌。乾隆皇帝的香妃是維吾爾族人,參照下圖, 應該是香的。
下圖是ABCC11 (rs17822931位點)的兩種等位基因(alleles)在全球人羣的分佈比例,藍色爲C(顯性溼耳垢),黃色爲T(隱性)。

From polyhedron 復旦大學現代人類學實驗室
人類走出熱帶非洲,在進入亞洲溫帶的過程中,第16號常染色體的ABCC11上發生基因突變,導致東方人的溼型耳垢(earwax)變乾, 還有母親的初乳分泌減少。 初乳(colostrum)是產仔後短時閒內分泌的乳汁,蛋白質含量比常乳高,包含有豐富的免疫球蛋白、乳鐵蛋白、生長因子等,有助於嬰兒增強體質。
溼性耳垢的人容易產生狐臭, 就是腋窩下出汗,分泌一些脂肪酸類, 這些本身氣味並不大,但被一些細菌分解, 產生一些臭的有機物。對於乾耳垢的人來說,因爲腋下脂肪酸分泌少,細菌也長得慢,哪怕幾天不洗澡也不會有狐臭。
但對於溼耳垢的人來說,在熱天只需要半天狐臭就會出現。東方人通常都是晚上洗澡,正好除去一天的灰塵睡覺。而歐洲人卻一般都是早起洗澡,如果早上不洗,晚上腋窩的汗液在上午也許就能讓細菌長起來而造成狐臭。
狐臭在全世界各人羣的分佈比例非常不同, 這個突變主要分佈在華北中原地區, 因此在中國,狐臭反而成了極少數。韓國人跟中國人差不多,但日本人當中只有84%沒有狐臭。大洋洲土著只有30%沒有狐臭,白人裡面只有10%沒有狐臭, 黑人中只有0.5%沒有狐臭。印第安人也是大多沒有狐臭,說明他們是從亞洲過來的。日本人有狐臭的人集中在阿伊努人中,說明與大和民族的不同起源。
在動物中,體味有很大作用,是吸引異性的信息素, 所以說有狐臭才是人的常態。中國古詩說女子“香汗薄衫涼”,一香一臭,看似相反,其實原理完全相同。少了淡了就香,多了濃了就臭,香奈兒5號打翻了瓶子也是要熏死人的。
中國自古就有“狐狸精”的傳說,可能與狐臭有關。在古代,西方女子皮膚白皙、身材好,同時不拘禮教,招東方男性的喜歡,自然引起東方女子的妒嫉, 認為她們是狐狸變的,所以有狐臭。狐臭本來稱作“胡臭”,是區別胡漢的一種標誌。乾隆皇帝的香妃是維吾爾族人,參照下圖, 應該是香的。
下圖是ABCC11 (rs17822931位點)的兩種等位基因(alleles)在全球人羣的分佈比例,藍色爲C(顯性溼耳垢),黃色爲T(隱性)。

From polyhedron 復旦大學現代人類學實驗室
Oct 16, 2015
匈奴、孔子、曹操的基因 Genes of Hunnu, Confucius & Emperor Cao
匈奴是什麼民族? 至今仍未能考証, 有許多爭議, 主要資料都是透過中國文獻。公元350年左右,匈人(Huns)進入了歐洲,他們和匈奴人是否有血緣關係, 尚無定論。近年來使用DNA測試也許可以回答這一問題。(請看下面圖表和解釋)
匈奴 = 蒙古語:Hunnu, 粵語:Hung1-nou4, 廣韻:Hiong-no, 上古漢語:Qhoŋ-naa
近年新疆哈密地區巴里坤-黑溝梁墓地的發掘, 被認為是漢代匈奴夏季王庭所在地, 墓地人類遺骸檢測出全是Q類型: Q1a*, Q1b, Q*。 後來的研究表明,還存在幾例Q-M3, 即印第安人的主要類型之一。
也是在匈奴影響區域, 寧夏彭陽縣出土東周時期古墓, 遺骸檢測出全是Q1a1類型, 是東亞常見的單倍型類群。 墓葬旁邊有青銅劍,春秋戰國祇有貴族才能佩劍。孔子即出身貴族。
山西絳縣橫水村2004年發現西周墓地, 失落3000年的倗國橫空出世, 不見於史籍記載, 也許在商時已經存在。 其葬俗有相當大的非周人傳統, 古DNA 證明Q1a1是當時貴族血統。
蒙古北部額金河 Egyin Gol 和 東北部 Duurlig Nars 發現2000年前的匈奴古墓,根據對古墓中的DNA檢測,基因類似目前北亞蒙古人種, 還有一例是R1a1: 印歐人種。請看下面圖表:

孔子自稱是殷人,身高191厘米,被後世稱為「長人」,這是父親的遺傳。 最近,研究人員對曲阜地區1118名孔姓男性進行DNA檢測,發現有3種高頻單倍群:C3,Q1a1和O3,雖然O3是通常漢族單倍型,前兩者有著明顯的單祖先擴散結構,最可能是孔子類型。 當然我們假設孔子的後代仍然在曲阜地區, 是這樣嗎?請看幾個歷史事件。
1. 孔末亂孔: 孔末是一個雜役,按當時僕隨主姓的習俗姓孔, 在五代十國時期,眼見天下大亂,時局動盪,起了謀逆奪位的野心,大殺孔子後裔及四十二世孫孔光嗣,儼然以孔子嫡裔自居。他的後代世稱「外孔」。
2. 孔仁玉中興: 聽說孔光嗣的獨生子孔仁玉剛滿九月,被母親張氏抱回娘家撫養成人。在後唐明宗長興元年,魯人將孔末告之於官府, 孔末被殺, 孔仁玉恢復嫡裔,但他也有可能是張氏的長子。他的後代世稱「內孔」。
3. 元朝滅南宋後,欲召南宗孔氏回曲阜襲封奉祀,孔洙辭讓,蒙古人另冊封一個北宗“衍聖公”, 於是山東曲阜的孔府就這麼傳下來了。
在2009年,河南省安陽市宣布發現了曹操墓,質疑者甚眾。2010 年復旦大學現代人類學實驗室推行項目研究,結論證明曹操是O2a型的, 應是父親從本宗室過繼, 而非抱養自街頭乞丐,請看下面圖表, 詳情觀看下面視頻。

請看視頻: Click to see 《Y染色體攜帶的歷史》 from polyhedron 復旦大學現代人類學實驗室
吳偉榮2015秋
匈奴 = 蒙古語:Hunnu, 粵語:Hung1-nou4, 廣韻:Hiong-no, 上古漢語:Qhoŋ-naa
近年新疆哈密地區巴里坤-黑溝梁墓地的發掘, 被認為是漢代匈奴夏季王庭所在地, 墓地人類遺骸檢測出全是Q類型: Q1a*, Q1b, Q*。 後來的研究表明,還存在幾例Q-M3, 即印第安人的主要類型之一。
也是在匈奴影響區域, 寧夏彭陽縣出土東周時期古墓, 遺骸檢測出全是Q1a1類型, 是東亞常見的單倍型類群。 墓葬旁邊有青銅劍,春秋戰國祇有貴族才能佩劍。孔子即出身貴族。
山西絳縣橫水村2004年發現西周墓地, 失落3000年的倗國橫空出世, 不見於史籍記載, 也許在商時已經存在。 其葬俗有相當大的非周人傳統, 古DNA 證明Q1a1是當時貴族血統。
蒙古北部額金河 Egyin Gol 和 東北部 Duurlig Nars 發現2000年前的匈奴古墓,根據對古墓中的DNA檢測,基因類似目前北亞蒙古人種, 還有一例是R1a1: 印歐人種。請看下面圖表:

孔子自稱是殷人,身高191厘米,被後世稱為「長人」,這是父親的遺傳。 最近,研究人員對曲阜地區1118名孔姓男性進行DNA檢測,發現有3種高頻單倍群:C3,Q1a1和O3,雖然O3是通常漢族單倍型,前兩者有著明顯的單祖先擴散結構,最可能是孔子類型。 當然我們假設孔子的後代仍然在曲阜地區, 是這樣嗎?請看幾個歷史事件。
1. 孔末亂孔: 孔末是一個雜役,按當時僕隨主姓的習俗姓孔, 在五代十國時期,眼見天下大亂,時局動盪,起了謀逆奪位的野心,大殺孔子後裔及四十二世孫孔光嗣,儼然以孔子嫡裔自居。他的後代世稱「外孔」。
2. 孔仁玉中興: 聽說孔光嗣的獨生子孔仁玉剛滿九月,被母親張氏抱回娘家撫養成人。在後唐明宗長興元年,魯人將孔末告之於官府, 孔末被殺, 孔仁玉恢復嫡裔,但他也有可能是張氏的長子。他的後代世稱「內孔」。
3. 元朝滅南宋後,欲召南宗孔氏回曲阜襲封奉祀,孔洙辭讓,蒙古人另冊封一個北宗“衍聖公”, 於是山東曲阜的孔府就這麼傳下來了。
在2009年,河南省安陽市宣布發現了曹操墓,質疑者甚眾。2010 年復旦大學現代人類學實驗室推行項目研究,結論證明曹操是O2a型的, 應是父親從本宗室過繼, 而非抱養自街頭乞丐,請看下面圖表, 詳情觀看下面視頻。

請看視頻: Click to see 《Y染色體攜帶的歷史》 from polyhedron 復旦大學現代人類學實驗室
吳偉榮2015秋
Aug 12, 2015
走入川西民族走廊, 尋找東女國 Ethnic Corridor of West Sichuan, Ancient female nation
川西有許多少數民族世世代代居住在青藏高原的東緣, 有著自己獨特的語言和文化, 他們的地理位置, 主要依據各族群的語言(包括正走向消亡的語言)來劃分。
這裡多數的族群在某種程度上與藏族生活方式認同,但也混合了自己獨特的傳統,有時也受到近鄰文化的影響。這裡大多屬羌支語言,不僅獨具特色,而且完全有別與藏語。
中國的少數民族定得很混亂,經常有講不同語支、完全不能互通的語言的族羣被分在同一民族,而講很相近的被分爲兩個民族。比如白馬人和嘉絨人,盡管在上世紀50年代他們都不覺得自己是藏人。還有比如瀘沽湖旁的摩梭人,語言較接近麗江的納西人,在雲南被劃爲納西族,而在四川被劃爲蒙古族。



上次談到白馬藏族是東亞最古老的部族,他們的祖先是氐族。現在談嘉絨藏族, 他們的祖先同樣不是藏族, 而是羌族。大渡河畔居民主要是嘉絨藏族(看上圖), 專家認為他們是古代「東女國」的後裔。據《舊唐書﹒南蠻西南蠻傳》記載:「東女國,西羌之別稱,以西海中復有女國,故稱東女焉。俗以女為王。」
嘉絨文化可被視為民族走廊之精髓:獨具特色的石頭樓房,經幡飛揚;古老的石砌碉樓,神奇叵測;更有那佩戴手織腰帶、裙飾和頭巾的羌族女人,花枝招展。據稱蒙古滅西夏國後,西夏大批后宮妃嬪從甘肅經川西高原流入丹巴一帶,定居於此,將美麗的血質注入這了一方。最近羌支語系人群(丹巴、道孚、新龍、雅江, 看上面地圖)有了DNA檢測, 下表是羌支語、藏、彝、漢族的單倍群表。

這裡請看視頻: 在甘孜州的首府康定城開始,隨著大渡河畔,來到美麗的丹巴, 乘坐大金川牛皮船, 尋找東女國。

Click to watch All videos on Youtube web page.
我另有Post觀賞川西旅遊。
這裡多數的族群在某種程度上與藏族生活方式認同,但也混合了自己獨特的傳統,有時也受到近鄰文化的影響。這裡大多屬羌支語言,不僅獨具特色,而且完全有別與藏語。
中國的少數民族定得很混亂,經常有講不同語支、完全不能互通的語言的族羣被分在同一民族,而講很相近的被分爲兩個民族。比如白馬人和嘉絨人,盡管在上世紀50年代他們都不覺得自己是藏人。還有比如瀘沽湖旁的摩梭人,語言較接近麗江的納西人,在雲南被劃爲納西族,而在四川被劃爲蒙古族。



上次談到白馬藏族是東亞最古老的部族,他們的祖先是氐族。現在談嘉絨藏族, 他們的祖先同樣不是藏族, 而是羌族。大渡河畔居民主要是嘉絨藏族(看上圖), 專家認為他們是古代「東女國」的後裔。據《舊唐書﹒南蠻西南蠻傳》記載:「東女國,西羌之別稱,以西海中復有女國,故稱東女焉。俗以女為王。」
嘉絨文化可被視為民族走廊之精髓:獨具特色的石頭樓房,經幡飛揚;古老的石砌碉樓,神奇叵測;更有那佩戴手織腰帶、裙飾和頭巾的羌族女人,花枝招展。據稱蒙古滅西夏國後,西夏大批后宮妃嬪從甘肅經川西高原流入丹巴一帶,定居於此,將美麗的血質注入這了一方。最近羌支語系人群(丹巴、道孚、新龍、雅江, 看上面地圖)有了DNA檢測, 下表是羌支語、藏、彝、漢族的單倍群表。

這裡請看視頻: 在甘孜州的首府康定城開始,隨著大渡河畔,來到美麗的丹巴, 乘坐大金川牛皮船, 尋找東女國。

Click to watch All videos on Youtube web page.
我另有Post觀賞川西旅遊。
Aug 8, 2015
青藏高原東緣的族群 Ethnic groups in eastern Tibetan Plateau
青藏高原東緣,是指北起甘肅南部、青海東部,經四川西部至雲南西北的弧形地域。
青海東北部是高原河谷,高度約在2000多公尺,大多是農區, 有藏族、土族、蒙古族、回族、撒拉族、漢族等。青海東邊接著甘肅南部的瑪曲、碌曲,地勢高為農牧混合區。再往東則是甘南的迭部、卓尼、舟曲等地,南接松潘之東的北川、平武。這些都是白龍江、湔江、涪江上游的高山縱谷地帶,居民主要是務農的藏、漢、回、羌族。青海東南到松潘草地是高原,有阿尼瑪卿與巴顏喀拉等山脈橫延其間,大部分地方皆只宜放牧, 居民主要是藏族。
松潘草原之南,東半部是岷江、大渡河及其支流切割而成的高山縱谷。河谷高度約在2000公尺上下,山巔則常超過4000公尺。由此往西是壤塘、爐霍、道孚、塔公等地,山勢一般較平緩,但河谷海拔更高,這是半農半牧區。這一帶鄉間住的主要是藏族,城鎮及其附近則有漢族、回族。
康定、瀘定及其以南的九龍、石棉仍是高山縱谷。同樣的,山谷是農區,愈往西河谷海拔愈高,牧業也愈重要。這兒鄉間住的主要是藏族、彝族,城鎮及其附近則有藏、彝、漢、回等族。在此之西,巴塘、理塘、九龍、稻城、鄉城等地河谷海拔更高,這是農、牧或半農半牧區。石棉以南,稻城、鄉城之東為安寧河流域。這兒有較廣闊的河谷平原,為務農的漢族、彝族所居。安寧河流域之東、南邊為涼山地區,雖仍為高山深谷,但一般來說河谷平原較寬,山邊坡地也較緩,這兒居住的大多為彝族。
青海東北部是高原河谷,高度約在2000多公尺,大多是農區, 有藏族、土族、蒙古族、回族、撒拉族、漢族等。青海東邊接著甘肅南部的瑪曲、碌曲,地勢高為農牧混合區。再往東則是甘南的迭部、卓尼、舟曲等地,南接松潘之東的北川、平武。這些都是白龍江、湔江、涪江上游的高山縱谷地帶,居民主要是務農的藏、漢、回、羌族。青海東南到松潘草地是高原,有阿尼瑪卿與巴顏喀拉等山脈橫延其間,大部分地方皆只宜放牧, 居民主要是藏族。
松潘草原之南,東半部是岷江、大渡河及其支流切割而成的高山縱谷。河谷高度約在2000公尺上下,山巔則常超過4000公尺。由此往西是壤塘、爐霍、道孚、塔公等地,山勢一般較平緩,但河谷海拔更高,這是半農半牧區。這一帶鄉間住的主要是藏族,城鎮及其附近則有漢族、回族。
康定、瀘定及其以南的九龍、石棉仍是高山縱谷。同樣的,山谷是農區,愈往西河谷海拔愈高,牧業也愈重要。這兒鄉間住的主要是藏族、彝族,城鎮及其附近則有藏、彝、漢、回等族。在此之西,巴塘、理塘、九龍、稻城、鄉城等地河谷海拔更高,這是農、牧或半農半牧區。石棉以南,稻城、鄉城之東為安寧河流域。這兒有較廣闊的河谷平原,為務農的漢族、彝族所居。安寧河流域之東、南邊為涼山地區,雖仍為高山深谷,但一般來說河谷平原較寬,山邊坡地也較緩,這兒居住的大多為彝族。
Apr 10, 2015
人類的單倍羣、遷徙、歷史 Human's haplogroups, migration & history
各位只要花100美元基因檢測,就可以測你的單倍群(Haplogroup)。除非是外星人, 如果得到一個奇怪的結果,不要驚訝,因為這只是一個祖先,你還有2 的N次方個祖先。 單倍群和人種不一定有因果關係的(網上常誤導), 但可以探索族群起源與移徙。1990年美國投資三十億美元的人類基因組計劃(Human Genome Project)正式啟動,成為一個宏大 的跨國項目, 其宗旨在於測定人類基因及其序列,破譯遺傳信息, 對生命科學、醫學有深遠的影響。
單倍群是一組類似的單倍型(Haplotype), 有一個共同的古老基因突變(mutation), 可用來標記數千、萬年前的祖先來源。在人類遺傳學中,最常被拿來研究的單倍群, 是Y染色體(Y-DNA)單倍群(父傳子)和線粒體DNA(mtDNA)單倍群(母傳女)。 在下面的進化樹圖中,全世界人的Y-DNA單倍羣編號是從A到T字母。所有非洲之外男性都有一個最近共祖,在他身上有一個基因突變標記M168(一路發)。非洲之外人數雖然很多,但從進化樹上看,都只屬於比較下游的支系。

在7萬多年前人類走出非洲,其中D和E型的人群可能是在六、七萬年前紅海附近分離。攜帶E型的人回到非洲,成為非洲西部的Negroid大黑人;而攜帶D型的人輾轉向東遷徙,成為東南亞的Negrito小黑人。兩種人的分佈相距如此遙遠,而Negroid人非常高大,往往超過180厘米,而Negrito人一般不會超過150厘米。D型廣泛分佈於藏族及周邊民族, 約1萬年前, 獨特的類型D2-M55出現在日本。
跨過紅海後的C和F型人群,演化成了不同的人種。 C型在五、六萬年前擴散到東亞、東南亞、澳大利亞、南太平洋島嶼,也被稱為澳大利亞人種。而F型則是歐亞和印第安人的祖先, 大約在三、四萬年前F型開始擴張,其下有G、H、I、J、L、T在歐亞大陸西部成為高加索人種。高加索人種雖然往往被稱為白人,但是膚色不一定很白。K型下有N、O、Q、R, 大約2萬年前,Q和R型人群來到了中亞,部分Q型繼續東遷,大約1.5萬年前跨過白令海峽進入美洲,形成印第安人種。 R型是中亞地區的主要類群,但同時大量向西遷徙加入高加索人種,成為歐洲常見人群。
也是大約2萬年前N和O型人群來到東亞。大約1.3萬年前,N型的人從東亞擴張到北亞和北歐。這部分人在歐洲號稱自己是黃種人,但怎麼看都不像,也許他們覺得自己跟周圍的白種人差別太大,把這些差異給放大了。然後O型是東亞人最主要的類型,在東亞人裡面,幾乎占70到80%, 下面分O1、O2、O3三大種類。 O1出現在中國東南的百越民族的後代,包括侗傣語係與南島語系的這些民族,台灣高山族像阿美族占100%。O2型分兩部分,一個分支出現在華南、南亞、東南亞,另外一個分支出現在日本、朝鮮、中國東北。中國大部分民族裡面都有O3,在漢族裡占到50–60%。
吳偉榮2015秋
單倍群是一組類似的單倍型(Haplotype), 有一個共同的古老基因突變(mutation), 可用來標記數千、萬年前的祖先來源。在人類遺傳學中,最常被拿來研究的單倍群, 是Y染色體(Y-DNA)單倍群(父傳子)和線粒體DNA(mtDNA)單倍群(母傳女)。 在下面的進化樹圖中,全世界人的Y-DNA單倍羣編號是從A到T字母。所有非洲之外男性都有一個最近共祖,在他身上有一個基因突變標記M168(一路發)。非洲之外人數雖然很多,但從進化樹上看,都只屬於比較下游的支系。

在7萬多年前人類走出非洲,其中D和E型的人群可能是在六、七萬年前紅海附近分離。攜帶E型的人回到非洲,成為非洲西部的Negroid大黑人;而攜帶D型的人輾轉向東遷徙,成為東南亞的Negrito小黑人。兩種人的分佈相距如此遙遠,而Negroid人非常高大,往往超過180厘米,而Negrito人一般不會超過150厘米。D型廣泛分佈於藏族及周邊民族, 約1萬年前, 獨特的類型D2-M55出現在日本。
跨過紅海後的C和F型人群,演化成了不同的人種。 C型在五、六萬年前擴散到東亞、東南亞、澳大利亞、南太平洋島嶼,也被稱為澳大利亞人種。而F型則是歐亞和印第安人的祖先, 大約在三、四萬年前F型開始擴張,其下有G、H、I、J、L、T在歐亞大陸西部成為高加索人種。高加索人種雖然往往被稱為白人,但是膚色不一定很白。K型下有N、O、Q、R, 大約2萬年前,Q和R型人群來到了中亞,部分Q型繼續東遷,大約1.5萬年前跨過白令海峽進入美洲,形成印第安人種。 R型是中亞地區的主要類群,但同時大量向西遷徙加入高加索人種,成為歐洲常見人群。
也是大約2萬年前N和O型人群來到東亞。大約1.3萬年前,N型的人從東亞擴張到北亞和北歐。這部分人在歐洲號稱自己是黃種人,但怎麼看都不像,也許他們覺得自己跟周圍的白種人差別太大,把這些差異給放大了。然後O型是東亞人最主要的類型,在東亞人裡面,幾乎占70到80%, 下面分O1、O2、O3三大種類。 O1出現在中國東南的百越民族的後代,包括侗傣語係與南島語系的這些民族,台灣高山族像阿美族占100%。O2型分兩部分,一個分支出現在華南、南亞、東南亞,另外一個分支出現在日本、朝鮮、中國東北。中國大部分民族裡面都有O3,在漢族裡占到50–60%。
吳偉榮2015秋
Apr 8, 2015
走出伊甸園, 羊的傳人 Out of Eden, Shepherd Linkage
在7萬多年前, 有幾百人在紅海南端走出非洲,他們就是全世界70億人類(不包括非洲裔)的祖先。我們是怎麼知道的? 在2005年, 有一項龐大的國際「基因地理計劃」( Genographic Project), 藉在全球收集人類的基因樣本,探索人類起源及全球移徙擴散,其中東亞和東南亞地區的研究, 是由復旦大學現代人類學實驗室負責的, 目前基本框架已經明晰了。
出走非洲有一位男性,他的Y染色體(父傳子)有一個基因突變標記,叫M168(中文:一路發),他是70億人的祖先, 其中全部男性都有這標記。(母傳女)是用線粒體mtDNA標記, 現代人的基因可以追溯到約十幾萬年前的一位非洲女性,科學家稱為「粒線體夏娃」(Mitochondrial Eve), 所有的人都是她的後代。與她相對的父系祖先「Y染色體亞當」(Y-chromosomal Adam),是生存在不同年代, 估計也超過十幾萬年。“亞當”和“夏娃“不必相遇,只要他們的基因分別流傳下來就夠了。
Click to see larger map:
 在7萬多年前, 地球處於末次冰期,氣候迅速變冷, 非洲大陸出現大範圍的乾旱, 使人類出走非洲, 直接通過阿拉伯半島、印度南部,沿海岸線遷徙到東南亞, 約5~6萬年前到達了澳大利亞(左圖左邊: C4群), 而漢藏苗瑤族的祖先, 要在2萬年前, 沿著青藏高原東邊向北遷徙, 才走入中國(左圖右邊: O3群)。
在7萬多年前, 地球處於末次冰期,氣候迅速變冷, 非洲大陸出現大範圍的乾旱, 使人類出走非洲, 直接通過阿拉伯半島、印度南部,沿海岸線遷徙到東南亞, 約5~6萬年前到達了澳大利亞(左圖左邊: C4群), 而漢藏苗瑤族的祖先, 要在2萬年前, 沿著青藏高原東邊向北遷徙, 才走入中國(左圖右邊: O3群)。
目前專家認為, O3人群到達了中國的西部, 形成古羌族群。 後來一些古羌人往東定居, 學會了農業種植,不再是遊牧民族,人口很快的增長,形成了兩大對立, 生活方式不同的漢人和羌人。 許多學者相信,秦周的祖先是古羌人, 商的祖先是從東北來的, 但這些民族後來都變成漢族了。我有另文從考古材料來看: 秦周商的不同。
“羌”字,最早就見於殷商甲骨文, “羊”和“人”的合文。商人有用羌人祭神的習俗, 在甲骨卜辭中,2000餘條有人祭, 其中記載“人牲”近8000為“羌”, 顯然商人是非常不同的民族。但學術界對“羌”是專指羌方國或羌族,還是商土以西各部族的泛稱,一直存在爭議。 漢人是後來的名稱, 源於漢朝, 華夏人名稱源於周朝, 我有另文討論: 華夏人是誰?
吳偉榮2015春
出走非洲有一位男性,他的Y染色體(父傳子)有一個基因突變標記,叫M168(中文:一路發),他是70億人的祖先, 其中全部男性都有這標記。(母傳女)是用線粒體mtDNA標記, 現代人的基因可以追溯到約十幾萬年前的一位非洲女性,科學家稱為「粒線體夏娃」(Mitochondrial Eve), 所有的人都是她的後代。與她相對的父系祖先「Y染色體亞當」(Y-chromosomal Adam),是生存在不同年代, 估計也超過十幾萬年。“亞當”和“夏娃“不必相遇,只要他們的基因分別流傳下來就夠了。
Click to see larger map:
在7萬多年前, 地球處於末次冰期,氣候迅速變冷, 非洲大陸出現大範圍的乾旱, 使人類出走非洲, 直接通過阿拉伯半島、印度南部,沿海岸線遷徙到東南亞, 約5~6萬年前到達了澳大利亞(左圖左邊: C4群), 而漢藏苗瑤族的祖先, 要在2萬年前, 沿著青藏高原東邊向北遷徙, 才走入中國(左圖右邊: O3群)。目前專家認為, O3人群到達了中國的西部, 形成古羌族群。 後來一些古羌人往東定居, 學會了農業種植,不再是遊牧民族,人口很快的增長,形成了兩大對立, 生活方式不同的漢人和羌人。 許多學者相信,秦周的祖先是古羌人, 商的祖先是從東北來的, 但這些民族後來都變成漢族了。我有另文從考古材料來看: 秦周商的不同。
“羌”字,最早就見於殷商甲骨文, “羊”和“人”的合文。商人有用羌人祭神的習俗, 在甲骨卜辭中,2000餘條有人祭, 其中記載“人牲”近8000為“羌”, 顯然商人是非常不同的民族。但學術界對“羌”是專指羌方國或羌族,還是商土以西各部族的泛稱,一直存在爭議。 漢人是後來的名稱, 源於漢朝, 華夏人名稱源於周朝, 我有另文討論: 華夏人是誰?
吳偉榮2015春
Apr 6, 2015
東亞民族單倍羣分佈 Haplogroup distribution in East Asia
從人類考古學來看,末次冰期發生於人類的舊石器時代與中石器時代, 隨著地球走出末次冰期,人類也進入了新石器時代(約1萬年前),開始從事農業和畜牧。最早到達東亞、東南亞的人群是C和D單倍群, G, J, N, O, Q和R, 都是末次冰期結束前分支出來的,很可能起源於東南亞。 C、D、N和O, 是東亞四個主要單倍群,約占到東亞男性的93%。 O是東亞最大的單倍群,約占75%的中國人以及超過50%的日本人。 O3是中國最常見的單倍群,遍及整個東亞和東南亞,佔漢族50-60%左右。 C、D、N、O和Q常見於歐亞大陸東部,J、G和R常見於歐亞西部。現在已知大部分的歐亞東部支系, 是在舊石器時代晚期產生的。(見下文人類遷徙Posts)
東亞各民族的分佈:(初步資料僅供參考)

漢族按方言區的分佈:(初步資料僅供參考)

幾乎全部的漢藏族群都有一個共同的Y染色體O3遺傳特徵,羌族的Y染色體多樣性在藏東裡是最高的。 如今漢藏緬語之間的關係已經非常清楚了,估算漢語與藏緬語約在6千年前分開。羌語支語言被認為是漢藏語系中最古老的類型,可能是漢藏語的源頭。
現在認為漢族、藏族、緬族、彝族、羌族、白族、景頗族、哈尼族、拉祜族、傈僳族、普米族、納西族、基諾族、德昂族、獨龍族、門巴族、珞巴族等源於古羌族。羌語支語言包括羌語、嘉戎語、爾龔語、拉塢戎語、普米語、木雅語、扎巴語、卻域語、貴瓊語、爾蘇語、史興語、納木依語等活著的語言及其文獻語言西夏語。
東亞各民族的分佈:(初步資料僅供參考)
漢族按方言區的分佈:(初步資料僅供參考)
幾乎全部的漢藏族群都有一個共同的Y染色體O3遺傳特徵,羌族的Y染色體多樣性在藏東裡是最高的。 如今漢藏緬語之間的關係已經非常清楚了,估算漢語與藏緬語約在6千年前分開。羌語支語言被認為是漢藏語系中最古老的類型,可能是漢藏語的源頭。
現在認為漢族、藏族、緬族、彝族、羌族、白族、景頗族、哈尼族、拉祜族、傈僳族、普米族、納西族、基諾族、德昂族、獨龍族、門巴族、珞巴族等源於古羌族。羌語支語言包括羌語、嘉戎語、爾龔語、拉塢戎語、普米語、木雅語、扎巴語、卻域語、貴瓊語、爾蘇語、史興語、納木依語等活著的語言及其文獻語言西夏語。
Apr 4, 2015
東亞、東南亞的人類遷徙 Human migration in East & Southeast Asia
Click to see larger map:
 人群的遷徙和分佈與氣候有著密切的關係,在距今約11萬至1萬之間,地球處於末次冰期, 海平面比較低,東南亞島嶼與大陸相連(叫巽他古陸Sundaland, 見左圖), 牛津大學教授Stephen Oppenheimer, 稱之為東方的伊甸園。 那時北方比較冷, 巽他古陸的天氣,食物和環境,則比較適合人類生活。
人群的遷徙和分佈與氣候有著密切的關係,在距今約11萬至1萬之間,地球處於末次冰期, 海平面比較低,東南亞島嶼與大陸相連(叫巽他古陸Sundaland, 見左圖), 牛津大學教授Stephen Oppenheimer, 稱之為東方的伊甸園。 那時北方比較冷, 巽他古陸的天氣,食物和環境,則比較適合人類生活。
距今2.65萬年到1.9-2萬年間(叫末次冰盛期),是氣候最寒冷、冰川規模最大時期,亞洲的絕大部分、北歐和北美都被冰雪覆蓋,人類的生存空間縮小。 大約1.5萬年前,氣溫開始轉暖,冰川也開始退卻,人類才迎來了遷徙的好時刻。
地球在氣溫開始轉暖時, 有一O3人群沿著青藏高原東邊向北遷徙,進入中國的西部, 形成了古羌族群, 或者稱為氐羌族群,而氐族與羌族是否為同一民族, 或者原先“氐”並非族稱, 至今未有定論。
現在許多羌語支語言的民族, 分佈在四川西部的河谷地區,這裡被稱為“四川民族(藏彝)走廊”,連接著黃河中上游和藏東,極可能是古人群起源和遷徙的通道。也有證據,有一支O3北遷徙進入中國東部地區。
Above map is from Wikipedia with possible migration routes according to the Coastal Migration Model (2013).
 人群的遷徙和分佈與氣候有著密切的關係,在距今約11萬至1萬之間,地球處於末次冰期, 海平面比較低,東南亞島嶼與大陸相連(叫巽他古陸Sundaland, 見左圖), 牛津大學教授Stephen Oppenheimer, 稱之為東方的伊甸園。 那時北方比較冷, 巽他古陸的天氣,食物和環境,則比較適合人類生活。
人群的遷徙和分佈與氣候有著密切的關係,在距今約11萬至1萬之間,地球處於末次冰期, 海平面比較低,東南亞島嶼與大陸相連(叫巽他古陸Sundaland, 見左圖), 牛津大學教授Stephen Oppenheimer, 稱之為東方的伊甸園。 那時北方比較冷, 巽他古陸的天氣,食物和環境,則比較適合人類生活。距今2.65萬年到1.9-2萬年間(叫末次冰盛期),是氣候最寒冷、冰川規模最大時期,亞洲的絕大部分、北歐和北美都被冰雪覆蓋,人類的生存空間縮小。 大約1.5萬年前,氣溫開始轉暖,冰川也開始退卻,人類才迎來了遷徙的好時刻。
地球在氣溫開始轉暖時, 有一O3人群沿著青藏高原東邊向北遷徙,進入中國的西部, 形成了古羌族群, 或者稱為氐羌族群,而氐族與羌族是否為同一民族, 或者原先“氐”並非族稱, 至今未有定論。
現在許多羌語支語言的民族, 分佈在四川西部的河谷地區,這裡被稱為“四川民族(藏彝)走廊”,連接著黃河中上游和藏東,極可能是古人群起源和遷徙的通道。也有證據,有一支O3北遷徙進入中國東部地區。
Above map is from Wikipedia with possible migration routes according to the Coastal Migration Model (2013).
Apr 2, 2015
走入藏區, 探秘東亞最古老的部族 East Asia's oldest tribe in Tibetan area
 事實上西藏只是藏區的一部分,傳統上藏區包括衛藏、安多和康區, 廣義的藏區還包含不丹、錫金與拉達克。「不丹」意思是「Bod的終結」,藏人自稱Bod, 暗示不丹是位於藏文化的最南端。
事實上西藏只是藏區的一部分,傳統上藏區包括衛藏、安多和康區, 廣義的藏區還包含不丹、錫金與拉達克。「不丹」意思是「Bod的終結」,藏人自稱Bod, 暗示不丹是位於藏文化的最南端。藏區古稱吐蕃, 現今讀"Tubo", 儘管以前的數千年中沒有任何記錄有這種讀法, 只有讀Tufan。另外日本人讀作Toban, 朝鮮人讀作Tupan, 越南人讀作Thổphồn, 都和漢語的讀音相映。
藏人可能來自古代西羌的一支“發羌”,“發”古音*pad,與bod音接近,很可能是其譯音。
另外白馬藏族並不是藏族,他們的祖先為氐人,現在發現他們是東亞最古老的部族。人類在7~8萬年前走出非洲,他們的祖先就在4~5萬年前到了中國,大約是人類到達澳大利亞時間,這是驚人的發現, 單倍群O型人在2萬年前,才走入中國。 氐與羌族是否為同一民族,下次再討論。(見前文Posts)
請看白馬藏族的基因型Genotype及東亞最古老的部族:

Click to watch All videos on Youtube web page.
Subscribe to:
Comments (Atom)